ИСХОДНОЕ СОСТОЯНИЕ
Исходное состояние натощак представляет собой переход от состояния сытости к состоянию голодания и может сохраняться в течение 6—12 ч после приема пищи (рис. 10—25). Хотя этот интервал и не отражает стабильного состояния, но тем не менее он является легко установимой точкой отсчета различных отклонений. В исходных условиях из жировой ткани высвобождаются жирные кислоты для удовлетворения энергетических потребностей мышц, сердца и паренхиматозных органов (печень, почки). Дыхательный коэффициент мышцы в покое близок к 0,7, что свидетельствует об окислении ею практически только жира. Углеводы утилизируются главным образом мозгом, в котором окисляется глюкоза до конца со скоростью 125 г/сут. Меньшие количества глюкозы утилизируются покоящейся мышцей и облигатно анаэробными тканями, такими, как форменные элементы крови и мозговой слой почек (рис. 10—26).
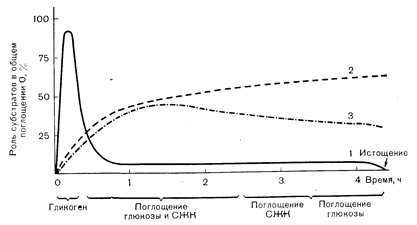
Рис. 10—25. Характер утилизации субстратов мышцами при физической нагрузке различной продолжительности. В первые несколько минут работы главным «топливом», используемым для образования АТФ, служит мышечный гликоген (1). Между 10-й и 40-й минутой работы по 40% всех окисляемых субстратов приходится на долю приносимых с кровью глюкозы и свободных жирных кислот (СЖК). Затем (т. е. при очень длительной работе) происходит прогрессивное увеличение утилизации СЖК (2) и снижение поглощения глюкозы (3) из крови. Хотя мышечный гликоген играет небольшую роль в качестве утилизируемого «топлива» при длительной работе, его полное истощение сопровождается развитием утомления.
Если в утилизации глюкозы принимают участие несколько тканей, то продукция ее ограничена печенью. Поддержание эугликемии зависит от высвобождения глюкозы печенью со скоростью, равной скорости ее утилизации в мозге и периферических тканях вместе (150—250 г/сут, 2—3 мг/кг в 1 мин, 6—10 г/ч). Примерно 15% глюкозы, высвобождаемой после ночного голодания, образуется из гликогена, остальное количество — при глюконеогенезе из лактата, аланина и в меньшей степени пирувата и глицерина [10]. При полном отсутствии продукции глюкозы в печени уровень сахара в крови должен был бы снизиться вдвое в течение 40—60 мин. Поступление жирных кислот в печень приводит к образованию кетоновых тел. Однако скорость кетогенеза в исходном состоянии может обеспечить только не превышающий 0,5 мМ уровень кетокислот в крови.
Гормональным сигналом, позволяющим начаться гликогенолизу, глюконеогенезу и кетогенезу, в исходном состоянии служит снижение уровня инсулина в плазме с 30—100 мкЕД/мл (в состоянии сытости) до 10—20 мкЕД/мл. Таким образом, утилизация
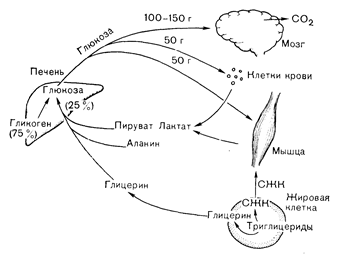
Рис. 10—26. Кругооборот глюкозы в состоянии покоя (после ночного голодания) у здорового человека. Единственным местом продукции глюкозы за счет гликогенолиза (75%) и глюконеогенеза (25%) является печень. Главным местом утилизации глюкозы, потребляющим ее со скоростью 100—150 г/сут, является мозг. Меньшие количества глюкозы поглощаются форменными элементами крови и покоящейся мышцей, служащей источником предшественников глюкозы в виде аланина и интермедиатов гликолиза—лактата и пирувата. Небольшие количества глицерина для глюконеогенеза поставляются адипоцитами. глюкозы в исходном состоянии не зависит от инсулина и почти полностью происходит в мозге. С другой стороны, сохранение исходного Уровня инсулина (и исходной скорости его секреции 0,25—1,5 ЕД/ч) удерживает продукцию глюкозы и кетоновых тел на уровне, не превышающем скорости их утилизации, что предотвращает развитие гипергликемии и гиперкетонемии. При уменьшении скорости секреции инсулина (например, при ювенильном диабете) или в условиях гормональных изменений, препятствующих действию исходного уровня инсулина (см. ниже, раздел «Стрессорная гипергликемия»), концентрация глюкозы и кетоновых тел в крови увеличивается.
Дата добавления: 2015-01-19; просмотров: 1247;