Nbsp; ПАТОГЕНЕЗ Т-КЛЕТОЧНОЙ ЛИМФОМЫ, ИНДУЦИРОВАННОЙ HTLV-1
Механизм индукции Т-клеточной лимфомы вирусом НТLV-1
(по V. Cumar, R. S. Cotran, S.L. Robbins, 1997)
· HTLV-1 не содержит v-onc и не способен к интеграции с протоонкогеном. Предполагается, что его трансформирующая активность связана с участком генома – tax. Продукт этого гена – белок tax активирует транскрипцию клеточных онкогенов (c -fos, c – sis), генов, кодирующих ИЛ - 2, рецепторы к ИЛ - 2, а также гранулоцитарно-макрофагальный колониестимулирующий фактор – GM - CSF.
· Поскольку HTLV-1 экспрессируют ИЛ-2 и рецепторы к нему вне зависимости от субклоновой принадлежности Т-клеток, последние постоянно стимулируются к пролиферации аутокринным и паракринным путем. Происходит пролиферация всех клонов Т-лимфоцитов, т.н. поликлональная пролиферация.
· Одновременно под влиянием вируса HTLV-1 в Т-лимфоцитах происходит активация генов, контролирующих продукцию гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF). Под действием последнего усиливается наработка клеток моноцитарно-макрофагального ряда и их активация.
· Активированные таким путем макрофаги продуцируют ИЛ-1, который оказывает дополнительное костимулирующее влияние на пролиферацию Т-лимфоцитов.
· Активно пролиферирующие Т-клетки характеризуются нестабильностью генома, определяющей высокий риск появления дополнительных мутаций.
· В случае, если такая мутация имеет место, а мутировавшая клетка не элиминируется, продолжая активно пролиферировать, происходит образование ее многочисленных потомков с измененным геномом, т.е. моноклона Т-клеточной лейкемии.
· Таким образом, моноклональная Т-клеточная лейкемия/лимфома есть результат пролиферации одной единственной Т-клетки, подвергшейся дополнительной мутации.
ВИРУСНЫЙ КАНЦЕРОГЕНЕЗ
Опухолеродные ДНК-содержащие вирусы
· Онкогены ДНК - содержащих вирусов являются необходимыми вирусными генами и в отличие от РНК - содержащих не имеют клеточных гомологов.
· В отличие от РНК - содержащих онкогенных вирусов, трансформирующий потенциал которых не связан с продукцией вирусных частиц, онкогенные свойства ДНК - содержащих вирусов тесно связаны со способностью этих вирусов к продуктивной инфекции.
· Инфекция клеток, которая является пермиссивной для репликации вируса, будет приводить к образованию значительного количества вирусных частиц и к гибели клетки. В этом случае трансформирующий потенциал вирусов не может быть выражен.
· Феномен трансформации может проявляться только в тех случаях, когда инфекция происходит в непермиссивных условиях, при которых процесс репликации вируса ограничен.
· Трансформирующий потенциал онкогенных ДНК - содержащих вирусов проявляется прежде всего в их способности стимулировать покоящиеся клетки к вхождению в клеточный цикл, т.е. к нарушению контроля клеточного деления. Этот процесс является следствием функционирования вирусных онкогенов.
· Вирусные инфекции делятся на раннюю и позднюю стадии. В каждой из этих стадий действуют разные вирусные гены. Ранние гены кодируют белки, участвующие в репликации вирусной ДНК. Поздние гены – регулируют синтез белков вирусной оболочки. Для ДНК - содержащих вирусов только ранние гены ответственны за трансформацию клеток.
· Общий принцип взаимодействия ДНК-содержащих вирусов с клетками предусматривает возможность существования вирусного ДНК в клетке в двух формах: в эписомальной или в интегрированной (в составе клеточного генома). При этом в любом случае это взаимодействие предусматривает экспрессию ранних вирусных (трансформирующих) генов.
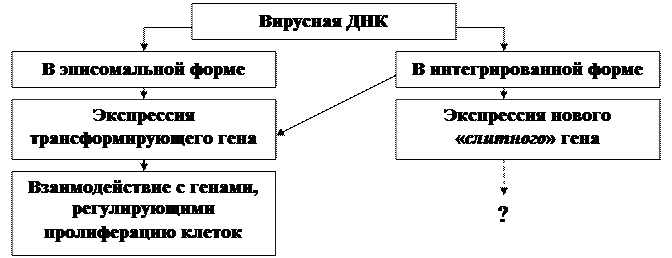

Общий принцип взаимодействия ДНК - содержащих вирусов с клетками
(Ф.Л. Киселев, 2004)
· Молекулярные механизмы трансформирующего действия этих ранних генов связаны с индукцией синтеза белковых продуктов, способных взаимодействовать с различными клеточными генами или кодируемыми ими белками, осуществляющими контроль за пролиферацией клеток.
| ДНК - содержащие вирусы | Вирусные гены, участвующие в трансформации | Внутриклеточные механизмы нарушения контроля клеточного цикла при экспрессии онкогенов |
| Вирус папиллом | Е 6 Е 7 | Инактивация гена - супрессора р53 Инактивация гена - супрессора р Rb105 Взаимодействие с циклинами |
| Вирус Эпштейна-Барра | EBNA-LP и EBNA 2 LMP и EBNA 3 | Взаимодействие с циклином D2 Взаимодействие с геном резистентности к интерферону Взаимодействие с геном bcl 2 |
| Вирус гепатита В | Х | Активация транскрипции собственных и других вирусных генов. Трансактивация некоторых факторов транскрипции |
· Возможна реализация еще одного механизма, связанного с интеграцией вирусного генома в клеточной, который может приводить к формированию нового «слитного» гена, содержащего как вирусную, так и клеточные последовательности с неизвестными функциями. Количество таких новых «слитных» генов невелико и их роль в канцерогенезе неизвестна.
· К ДНК-содержащим онкогенным вирусам относятся: вирусы группы Папова (папиллом, полиомы, вакуолизирующий вирус обезьян SV-40), аденовирусы, герпесвирусы и др.
· Из патогенных для людей наиболее изучены вирусы папиллом человека (HPV), Эпштейна-Барра (EBV) и вирус гепатита В (HBV).
ВИРУСНЫЙ КАНЦЕРОГЕНЕЗ
Опухолеродные ДНК-содержащие вирусы
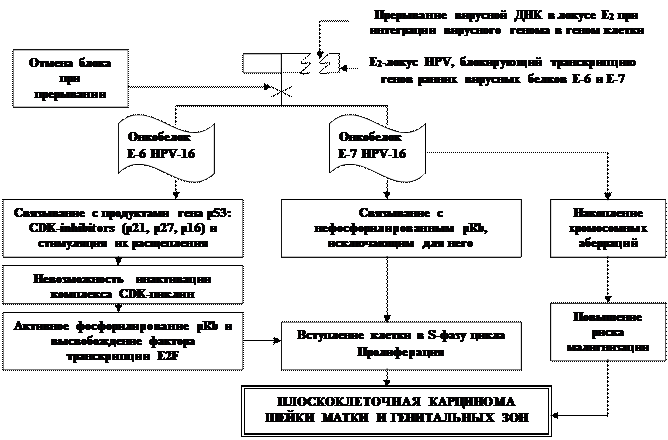 |
Молекулярные механизмы канцерогенного действия вируса папилломы человека
(по Ф.Л. Киселеву, 2004)
· Трансформирующие ДНК-содержащие вирусы папиллом (HPV-16; HPV-18) при реализации канцерогенного действия формируют стабильный комплекс с клеточным геномом хозяина. Если они поддерживаются в клетке в эписомальной (неинтегрированной) форме, развивается дисплазия и папилломы.
· Сайты интеграции ДНК HPV-16 (18), обнаруживаемых у 85% больных инвазивным плоскоклеточным раком шейки матки и карциномы in situ, с ДНК клетки случайны и определяются в разных участках генома хозяина.
· При интеграции происходит разрыв в вирусном геноме, который необходим для встраивания последнего в геном хозяина. Разрыв осуществляется, в основном, в рамках Е1 – Е2 – локусов вирусного генома.
· В результате интегрированный вирус утрачивает способность к завершению своего репликационного цикла, поскольку последовательности вирусных генов, необходимый для такого завершения, прерваны в ходе интеграции вирусной ДНК в геном хозяина.
· В то же время происходит разблокирование встроенных ранних вирусных генов Е-6 и Е-7, кодирующих транскрипцию соответствующих онкобелков.
· Влияние этих онкобелков на механизмы контроля клеточного цикла и обусловливают возможность злокачественной трансформации клеток.
Дата добавления: 2015-03-26; просмотров: 2218;