ПРОТООНКОГЕНЫ. ОНКОГЕНЫ. ОНКОСУПРЕССОРЫ.
ОПРЕДЕЛЕНИЕ ПОНЯТИЙ
(продолжение)
· Протоонкогены, в том числе гомологичные вирусным онкогенам, могут активироваться и без участия вирусов в результате процессов хромосомных транслокаций, амплификаций, мутаций.
· В нормальных клетках присутствуют белковые факторы негативной регуляции клеточного деления. Кодирующие их гены носят название генов-супрессоров (Rb, р53, АРС и др.). Реализация эффектов онкогенов находится в прямой зависимости от супрессии механизмов негативной регуляции клеточного деления.
· Таким образом, полная трасформация клетки является многоступенчатым процессом для достижения которого необходимо несколько генетических событий: активации онкогенов и инактивации генов, осуществляющих супрессорную функцию. При этом разные онкобелки действуют по общим биохимическим путям, в связи с чем разные онкогены могут дополнять эффекты друг друга, но один и тот же онкоген способен вызвать разные эффекты в клетках различного гистогенеза.
· Непрерывность роста трансформированных клеток поддерживается их нечувствительностью к апоптозу вследствие угнетения продукции проапоптотических факторов и усиления продукции антиапоптотических факторов:
 |
Раздел IV.
ПРОИСХОЖДЕНИЕ И МЕХАНИЗМЫ АКТИВАЦИИ ОНКОГЕНОВ
 |
(Г.И. Абелев, 1997)
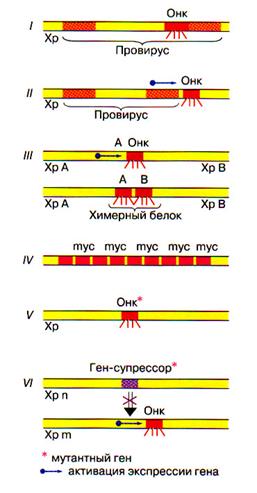
· I - вирусный онкоген: часть вирусного генома, интегрированного в хромосому хозяина (провирус). Онкогены ретровирусов: src, myc, ras, erb;
· II - активация клеточного онкогена встроенным провирусом (онкоген Int-1 при раке молочных желез мышей);
· III - хромосомные транслокации - разрыв и воссоединение фрагментов разных хромосом в единую новую хромосому могут привести к активации онкогена или образованию нового онкогена. В первом случае молчащий регуляторный ген попадает под контроль работающего клеточного гена и сам активируется (ген myc при лимфоме Бeркитта); во втором случае в месте разрыва - воссоединения образуется новый химерный ген, ведущий к синтезу химерного белка (белок bcr-abl при хроническом миелолейкозе);
· IV - амплификация (увеличение числа копий протоонкогена), ведущая к опухолевой трансформации (амплификация гена myc при опухолях нервной системы);
· V - мутация протоонкогена - синтез мутантного онкобелка (с-ras, при спонтанных и канцерогенных опухолях);
· VI - инактивация или утрата гена-супрессора опухолевого роста, ведущая к активации онкогена): ген Rb при ретинобластоме человека; ген р53 при разных опухолях человека).
ХРОМОСОМНЫЕ ТРАНСЛОКАЦИИ И СВЯЗАННЫЕ С НИМИ ОНКОГЕНЫ ХРОНИЧЕСКОЙ МИЕЛОИДНОЙ ЛЕЙКЕМИИ
 |
Хромосомные транслокации в развитии хронического миелолейкоза
(V. Cumar, R.S. Cotran, S.L. Robbins, 1997)
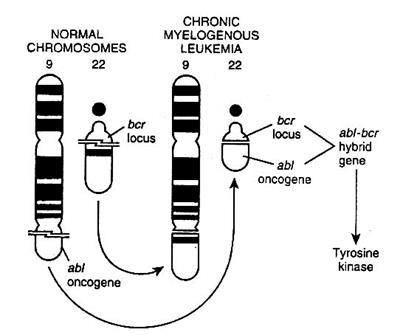
· На хромосоме 9 у человека картирован онкоген abl, являющийся эквивалентом онкогена вирусной лейкемии мышей Абельсона (v-abl). Он участвует в регуляции клеточного цикла и апоптоза.
· На хромосоме 22 располагается генетический локус bcr, кодирующий белок-рецептор bcr, воспринимающий внешние сигналы к развитию апоптоза.
· При реципрокной транслокации между хромосомами 9 и 22, онкоген abl встраивается в хромосому 22 в участке разрыва локуса bcr. В результате формируется аномальная т.н. филадельфийская хромосома с укороченным длинным плечом (Ph+), а из гена abl и локуса bcr образуется гибридный ген abl/bcr.
· Данный ген экспрессируется и кодирует синтез белка р-210, обладающего тирозинкиназной активностью. Поскольку тирозинкиназы - важная составляющая в передаче сигналов, в том числе ростовых факторов, синтез белка р-210 приводит к безудержной пролиферации клеток (апоптоз при этом подавляется) и к развитию хронического миелоидного лейкоза.
· Присутствие Ph+ не только в трансформированных клетках, но также во всех клетках миелоидного ростка кроветворения, включая гранулоцитарный, мегакариоцитарный и эритроидный ростки, служит неоспоримым доказательством происхождения хронического миелоидного лейкоза из одного патологического клона, родоначальницей которого является плюрипотентная стволовая клетка предшественница миелопоэза (КоЕ-ГЭММ).
· Несмотря на то, что химерный ген abl /bcr присутствует во всех ростках костного мозга, безудержный рост в развернутую стадию хронического миелоидного лейкоза касается, как правило, одного гранулоцитарного ростка. Реже имеет место повышенная выработка мегакариоцитов.
 |
ХРОМОСОМНЫЕ ТРАНСЛОКАЦИИ И СВЯЗАННЫЕ С НИМИ ОНКОГЕНЫ ЛИМФОМЫ БЕРКИТТА
Хромосомные транслокации в развитии лимфомы Беркитта
(V. Cumar, R. S. Cotran, S. L. Robbins, 1997)
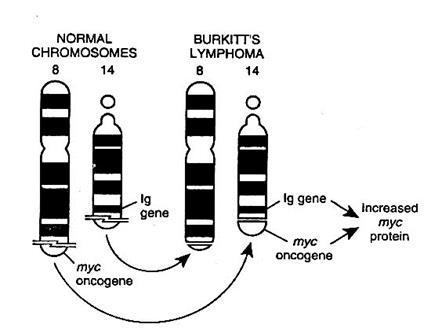
· На хромосоме 8, постоянно участвующей при всех типах транслокации при лимфоме Беркитта, картирован регуляторный онкоген myc, который является эквивалентом онкогена v-myc вируса миелоцитоматоза птиц (MYC). Пока этот онкоген (myc) располагается на 8 хромосоме, он репрессируется, т.е. представляет из себя "молчащий" онкоген.
· В В-лимфоцитах активно работают гены иммуноглобулинов. Так, в хромосоме 2 располагается локус, кодирующий синтез легкой цепи каппа (c); в хромосоме 22 - локус, кодирующий синтез легкой цепи лямбда (l), а в хромосоме 14 - гены, кодирующие синтез тяжелых цепей. Т.о., генетические локусы, контролирующие синтез тяжелых и легких цепей иммуноглобулинов, локализованы раздельно и даже в разных хромосомах.
· При транслокации гена myc с 8 хромосомы на 2, или с 8 на 22 хромосому он располагается рядом с генетическими локусами, кодирующими синтез легких цепей. При перемещении гена myc с 8 хромосомы на 14, он транслоцируется к иммуноглобулиновому локусу тяжелых цепей. В последнем варианте транслокация (между 8 и 14 хромосомами) носит реципрокный (взаимный) характер.
· В любом случае, после подобных транслокаций уровень экспрессии гена myc аффектируется соседними генами иммуноглобулинов, которые, в свою очередь, подпадают под контроль гена myc. Таким образом, нормальный клеточный контроль утрачивается.
· Экспрессия гена myc обусловливает безудержный рост клетки (В-лимфоцита), попавшей под влияние онкогена (лимфома Беркитта). При этом, в зависимости от того, на какую из хромосом произошла транслокация myc, клетки лимфомы синтезируют моноклональные иммуноглобулины с определенной характеристикой: легкой (c или l) или тяжелой (m, g, a, x, s) цепи.
Дата добавления: 2015-03-26; просмотров: 3156;