Роговая оболочка 2 страница
Роговая оболочка и склера
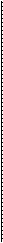 | |||
 | |||
|
| Роговица |
| Склера |
| 3D О О О О О О ОО 00 О О О 00 О О О ОО |
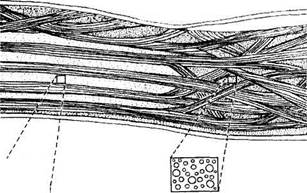
Рис. 3.2.8. Особенности расположения и взаимоотношения коллагеновых пластин роговой оболочки и склеры. Обращает на себя внимание различный диаметр коллагеновых волокон, расположенных в склере (по Bron et al., 1997)
ценна и отражает особенности архитектоники распределения коллагеновых пластин передних слоев стромы.
Параллельное расположение пластин передних отделов стромы роговицы и сохранение подобного расположения на границе с задними слоями позволяют производить межпластинчатое расслоение роговой оболочки при кератопластике [713].
Необходимо отметить, что передние и задние слои стромы отличаются как строением, так и физико-химическими свойствами. Так, задние слои стромы более упорядочены [343], более гидратированы [1117], обладают более низким преломляющим индексом [818]. Кроме того, коллагеновые пластины задних слоев стромы шире и толще (100—200 мкм— ширина и 1,0—2,5 мкм — толщина) передних слоев (0,5—30 мкм — ширина и 0,2—1,2 мкм — толщина) [603, 762, 763]. Имеются также и определенные различия строения кератоцитов [838].
Существование структурных различий передних и задних слоев стромы роговицы многие авторы рассматривают как основу большей устойчивости передних слоев к отеку. Именно это свойство обеспечивает сохранение кривизны роговицы и ее прозрачность при различных физиологических и патологических состояниях [764].
Стромальные пластины погружены в основное вещество, представленное различными типами протеогликанов. Гидрофильная часть основного вещества гликозаминогликанов, в которую погружены коллагеновые волокна, приобретает форму протеогликанов путем кова-лентного соединения гликозаминогликанов с белками. Протеогликаны имеют довольно разнообразное химическое строение. В строме роговой оболочки из гликозаминогликанов обнаружены кератан сульфат, хондроитин-4-суль-
фат, ходроитин-6-сульфат, дерматан сульфат [33, 34, 195].
Молекулы гликозаминогликанов окутывают волокна и ориентируются перпендикулярно кол-лагеновому волокну. Именно связь между волокнами и протеогликанами опеспечивает прозрачность роговичной ткани [983].
Различные типы гликозаминогликанов в роговой оболочке распределены неравномерно. Некоторые из них преобладают в передних слоях стромы, другие — в задних слоях. С преобладанием того или иного типа гликозаминогликанов в различных слоях стромы связана различная степень гидратации стромы [132, 579], с которой частично связана прозрачность стромы. Нарушение синтеза гликозаминогликанов (врожденное или приобретенное) приводит к помутнению роговицы, связанному с отложением продуктов патологического синтеза.
Клетки стромы (кератоциты).Основным клеточным элементом стромы роговой оболочки является кератоцит. Кератоциты составляют 2,4—5,0% объема стромы.
Наиболее близки кератоциты по происхождению и строению к фиброцитам. Обнаруживаются они во всех участках стромы, но с различной плотностью. Использование конфокальной микроскопии позволило установить, что плотность кератоцитов в центральных участках роговой оболочки равняется 20,5 ±2,9 кл/мм3. Отмечено также, что в передних слоях стромы их плотность меньше на 10%. Плотность кератоцитов уменьшается с возрастом примерно на 0,45% в год [817].
Кератоциты обладают длинными отростками, ориентированными параллельно коллагено-вым пластинам. Контактируют отростки с отростками рядом расположенных клеток этого же уровня, а также и клетками других уровней стромы (рис. 3.2.3, 3.2.7). При этом между ними формируются межклеточные контакты типа щелевых контактов [1151]. Предполагают, что эти контакты служат взаимодействию между кератоцитами, расположенными в виде сети во всей строме роговицы.
Толщина кератоцитов равна примерно 2 мкм. При этом ядро выглядит непропорционально большим.
Иммуноморфологически в цитоплазме клеток выявлены коллагены III, V и VI типов [695, 698, 878].
Цитоплазма кератоцитов бедна органоидами. В прямом контакте с цитоплазматической мембраной можно обнаружить пятна базальнопо-добного волокнистого материала, особенно по периферии роговицы. Плотный контакт этого материала с коллагеновыми фибриллами стромы приводит к образованию периодической структуры. Вокруг многих кератоцитов отмечается скопление фибриллярного и зернистого материала, являющегося структурным компонентом будущих коллагеновых волокон и основ-
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 ного вещества. Кератоциты обладают большой степенью подвижности.
ного вещества. Кератоциты обладают большой степенью подвижности.
Основная функция кератоцитов — синтез межклеточного вещества и коллагеновых фибрилл в период эмбриогенеза, после повреждения роговицы, а также поддержание метаболизма стромы на протяжении всей жизни.
Birk и Trelstad [122] установили, что поверхность фибробластов отвечает за пространственную ориентацию коллагеновых фибрилл. Именно благодаря этому свойству формируются пучки.
В связи с тем, что метаболическая активность кератоцитов в норме снижена, эндоплаз-матическая сеть клеток развита слабо. Лишь после травмы и воспалительных изменений роговицы эндоплазматическая сеть становится хорошо заметной [628].
В строме роговицы встречаются лимфоциты, макрофаги и полиморфноядерные лейкоциты.
Задняя пограничная (десцеметова) пластинка(lamina limitans postrior corneae; Dece-mett). Десцеметова оболочка при световой микроскопии выглядит бесструктурной мембраной, покрывающей заднюю поверхность стромы роговицы (рис. 3.2.3, 3.2.9). В гистогенетическом и структурном смыслах она представляет собой базальную мембрану заднего эпителия роговицы (эндотелия), который ее и продуцирует. Эластичность является одной из наиболее важных ее характеристик. Волокна десцеметовой мембраны образуются на протяжении всей жизни человека. Толщина их при рождении равняется 3 мкм, а в старости — 8—12 мкм [540, 878].
Как и другие базальные мембраны, десцеметова оболочка PAS-положительна и состоит из коротких и тонких фибрилл (10 нм). Фибриллы, в свою очередь, образованы коллаге-
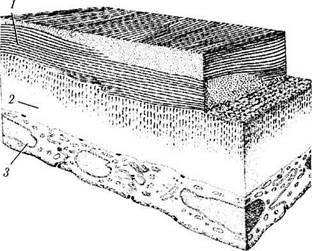
Рис. 3.2.9. Схема микроскопического строения задних слоев роговой оболочки (по Pouliquen, 1969):
1—строма роговой оболочки; 2—десцеметова оболочка; 3— задний эпителий (эндотелий)
ном IVтипа и погружены в гликопротеиновое основное вещество [316].
При ультраструктурном исследовании в мембране различают две области [98, 420, 496, 587]. Передняя ее треть имеет толщину 1—4 мкм и задние две трети — 5—15 мкм.
Передний слой десцеметовой оболочки, контактирующий со стромой, имеет многослойный пластинчатый вид, а задний —- гранулированный. Именно передний слой возникает в эмбриональном периоде первым. На тангенциальных срезах этот слой состоит из однородных пластин коллагеновых волокон, образующих равносторонние треугольники. Длина каждой стороны равна ПО нм. Треугольники связаны элек-тронноплотными узлами [1102]. Эти соединения появляются на 5 месяце внутриутробной жизни, когда слой имеют толщину 3,1 мкм (2,2 — 4,5 мкм). Задние 2/з мембраны образуются уже после рождения и состоят из гомогенного фиб-рогранулярного материала.
В мембране, помимо преобладающего коллагена IV типа, обнаружены коллагены III,V, VI и VIIIтипов [878].
С возрастом в десцеметовой мембране появляются, а затем увеличиваются в количестве коллагеновые волокна и слоистый материал. Этот процесс приводит к появлению на задней поверхности роговицы так называемых бородавок Хассал—Хенле (Hassal—Henle). При этом отмечается нарушение контактов между клетками эндотелия и нарушается барьерная функция последнего.
Несмотря на отсутствие в мембране Десце-мета эластических волокон, она исключительно эластична. При травме нередко десцеметова оболочка скручивается в виде рулона, что обнаруживается при биомикроскопии. Десцеметова мембрана исключительно устойчива в отношении протеолитических ферментов.
Эндотелий (задний эпителий роговой оболочки).Эндотелий роговой оболочки представляет собой один слой плоских гексагональных клеток (плоский однослойный эпителий), расположенных на десцеметовой оболочке (рис. 3.2.3, 3.2.9—3.2.11). Наиболее распространено мнение о том, что они происходят из клеток ней-рального гребня [792, 878, 1105].
Эндотелий роговой оболочки рассматривают как один из наиболее важных структурных компонентов, обеспечивающих прозрачность роговицы [451, 1145]. При этом показано, что обеспечение прозрачности роговицы связано со структурной организацией самой клетки, характера межклеточных контактов и расположением эндотелиальных клеток [128, 260, 261]. Основной функцией эндотелиальных клеток при этом является поддержание постоянного гидростатического давления стромы роговой оболочки. Именно важная роль эндотелия в сохранении прозрачности роговицы явилась причиной многочисленных исследований, направ-
Роговая оболочка и склера

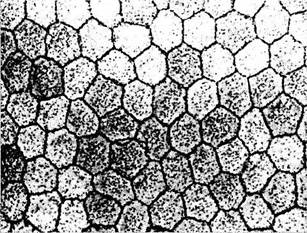
Рис. 3.2.10. Плоскостной препарат эндотелия центральных участков роговой оболочки при исследовании его в фазово-контрастном микроскопе:
отмечается полигональная форма клеток, их примерно одинаковые размеры и наличие плотных контактов между ними
ленных на изучение строения и функции этой структуры глаза. Способствовало этому применение эндотелиальной прижизненной микроскопии.
Последние исследования показали, что у взрослых количество эндотелиальных клеток ограничено и довольно постоянно. Их количество порядка 500 000. С возрастом число клеток уменьшается. Наибольшее уменьшение плотности эндотелиальных клеток определяется в первые годы жизни и полностью коррелирует с увеличением площади роговой оболочки ребенка.
При использовании эндотелиальной микроскопии установлено, что плотность эндотелиальных клеток при рождении колеблется в довольно широких пределах (2627—5316 клеток в мм2) [764]. Плотность клеток падает примерно на 26% на первом году жизни. Дальнейшее падение плотности клеток на 26% отмечается на протяжении последующих 2 лет. Затем скорость уменьшения плотности клеток снижается и число клеток стабилизируется к среднему возрасту [127, 767, 1001]. Кривая, отражающая процесс уменьшения плотности клеток, имеет линейную или логарифмическую форму [262].
В процессе дифференциации уменьшается степень полиморфизма эндотелиального пласта, а также уменьшается количество клеток гексаганальной формы [177, 259, 260, 262, 573, 765, 1113]. Правда, необходимо отметить, что скорость уменьшения плотности и формы клеток колеблется в широких пределах и не дает исследователям возможности сделать окончательное заключение относительно значения этого процесса и факторов, влияющих на этот процесс [1001, 1025].
У молодых людей размер клеток равен 18— 20 мкм (высота — 5—6 мкм), а в более позд-
нем возрасте — 40 мкм [1000]. Появляется би-модальность распределения клеток, как по размерам, так и по содержанию ДНК ядер [36].
Эндотелиальные клетки роговой оболочки присоединяются к десцеметовой оболочке при помощи полудесмосом. Рядом лежащие клетки плотно прилежат друг к другу и соединены десмосомами и запирательными пластинками. Запирательные пластинки распространяются по окружности апикальной поверхности клеток и закрывают межклеточные пространства, обеспечивая барьерные функции эндотелия. Рядом лежащие клетки соединяются также и при помощи «пальцевых вдавлений», представляющих собой цитоплазматические выросты, вдавливающиеся в тело соседней клетки. Несмотря на обилие межклеточных контактов, между клетками существуют щелевидные пространства, шириной 20 нм [163, 487].
Наличие контактов между клетками предопределяет пропускную способность эндотелиального слоя. Они ограничивают пассивный транспорт в строму роговой оболочки. Любое проникновение жидкости в строму через межклеточные щели уравновешивается активным ионным транспортом, происходящим трансцел-люлярно. Процессы регуляции проникновения жидкости в строму могут быть нарушены при уменьшении плотности расположения эндоте-лиоцитов и расширении межклеточных пространств, что нередко наблюдается при патологических состояниях. Рядом исследователей установлены также критические пределы уменьшения количества эндотелиальных клеток. Резкое уменьшение плотности клеток приводит к необратимому нарушению гидратации стромы. Считается, что такой плотностью клеток является величина, равная 400—700 клеток в квадратном миллиметре (при норме 1400— 2500 клеток) [578]. Тем не менее клинические наблюдения показывают, что при ряде патологических состояний даже существенное снижение плотности клеток далеко не всегда сопровождается усилением гидратации стромы роговицы [21].
На апикальной поверхности каждой эндотелиальной клетки располагается от 20 до 30 микроворсинок высотой 0,5—0,6 мкм и шириной 0,1—0,2 мкм. Именно эти образования значительно увеличивают площадь контакта клеточной поверхности с влагой передней камеры глаза. Можно обнаружить и реснички. Они чаще видны по периферии роговицы [889, 918]. Обнаружение ресничек позволило Hogan, Alva-rado, Weddell [496] предположить, что эндотелиальные клетки имеют единое происхождение с клетками трабекулярной сети.
Цитоплазма эндотелиоцитов богата митохондриями, которые обеспечивают энергией активный транспорт, секрецию и высокий уровень синтеза протеинов. Эндотелиоциты содержат митохондрии в значительно большем коли-
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 честве, чем любые другие клетки глаза за исключением рецепторных клеток. Обнаруживаются хорошо развитый гранулярный и аграну-лярный эндоплазматический ретикулум, многочисленные свободные рибосомы. Вблизи ядра четко виден аппарат Гольджи. Центриоли с ресничками располагаются в апикальной части клеток. В большом количестве определяются лизосомы. Отличительной чертой эндотелиаль-ных клеток является наличие многочисленных пиноцитозных пузырьков, связанных с цито-плазматической мембраной (рис. 3.2.11). Экспериментальными исследованиями с использованием радиоактивной метки показано быстрое перемещение этих пузырьков через цитоплазму в сторону десцеметовой мембраны. Иммуно-гистохимически в цитоплазме эндотелиальных клеток выявлены основные гликозаминоглика-ны роговицы — хондроитин-6-сульфат, хондрои-тин-4-сульфат, гепаран-сульфат.
честве, чем любые другие клетки глаза за исключением рецепторных клеток. Обнаруживаются хорошо развитый гранулярный и аграну-лярный эндоплазматический ретикулум, многочисленные свободные рибосомы. Вблизи ядра четко виден аппарат Гольджи. Центриоли с ресничками располагаются в апикальной части клеток. В большом количестве определяются лизосомы. Отличительной чертой эндотелиаль-ных клеток является наличие многочисленных пиноцитозных пузырьков, связанных с цито-плазматической мембраной (рис. 3.2.11). Экспериментальными исследованиями с использованием радиоактивной метки показано быстрое перемещение этих пузырьков через цитоплазму в сторону десцеметовой мембраны. Иммуно-гистохимически в цитоплазме эндотелиальных клеток выявлены основные гликозаминоглика-ны роговицы — хондроитин-6-сульфат, хондрои-тин-4-сульфат, гепаран-сульфат.
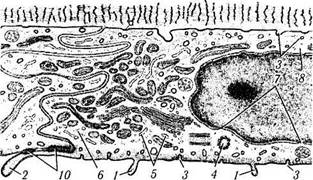
Рис. 3.2.11. Схематическое изображение ультраструктурной организации клеток эндотелия роговой оболочки:
/— микроворсинки; 2 — краевые выпячивания цитоплазмы в переднюю камеру глаза в местах межклеточных контактов; 3 — пиноцитозные пузырьки; 4 — центриоли; 5 — шероховатый эндоплазматический ретикулум; 6 — рибосомы; 7—ядерные поры; 8 — внутрицитоплазматические филаменты; 9 — аппарат Гольджи; 10 — межклеточные контакты различного типа
Необходимо остановиться и на основных физиологических функциях эндотелия роговицы. Одной из них является обеспечение клеток стромы питательными веществами. Процесс транспорта питательных веществ обеспечивается или диффузией между эндотелиоцитами, или активным переносом через содержимое клетки в направлении стромы.
Эндотелий играет главную роль в поддержании прозрачности роговицы путем активной регуляции содержания в строме воды. Эту функцию он выполняет, используя два механизма. Во-первых, он является активным барьером для солей и ряда метаболитов, проникновение которых в строму приводит к отеку последней. Во-вторых, он активно снижает осмотическое давление стромы наличием так называемого би-карбонатного насоса, возвращающего ионы из
стромы назад в камерную влагу [318, 711, 746, 918, 1204].
Кровоснабжение и лимфатическое дренированиероговицы обеспечиваются конъюнкти-вальными, эписклеральными и склеральными сосудами, являющимися ветвями передних ресничных артерий.
Нервы роговицы.Эпителий роговицы относится к наиболее интенсивно иннервируемым структурам организма человека. Чувствительная иннервация роговицы в 300—600 раз выше, чем иннервация кожи. Площадь эпителиального пласта, равная 0,01 мм2, содержит до 100 нервных окончаний [931]. На 2,1 млн ба-зальных клеток эпителия роговицы приходится до 1,4 млн нервных окончаний.
Сенсорная иннервация обеспечивается, в первую очередь, ветвями глазного нерва (ветвь тройничного нерва) [30, 878]. Главный источник иннервации роговицы — длинные ресничные нервы, являющиеся ветвями тройничного нерва.
Задний длинный ресничный нерв входит в склеру у заднего полюса и распространяется кпереди в супрахориоидальном пространстве. Различаются три уровня проникновения нервных окончаний в роговицу: склеральный, эпи-склеральный и конъюнктивальный [1222].
Около 80 нервных стволов проникают в ткань склеры вблизи лимба и, распространившись на 1—2 мм, теряют свои миелиновые оболочки. Эти волокна, покинув склеру, распределяются в средней трети стромы, делясь при этом ди- и трихотомически. Формируется в результате этого прекорнеальное сплетение. По мере продвижения к центральным участкам роговицы количество аксонов увеличивается за счет их последующего деления (рис. 3.2.12). При прохождении в строме роговой оболочки немиелинизованные нервные волокна располагаются параллельно коллагеновым пластинам. Отдельные нервные веточки подходят к керато-цитам и вдавливаются в их цитоплазматичес-кую оболочку [762, 763]. Окружают нервные стволы шванновские клетки и аморфный материал. Содержат аксоны многочисленные митохондрии, частицы гликогена и микропузырьки. Диаметр аксонов нервных волокон роговицы колеблется от 1 до 5 мкм.
В эпителиальный пласт из стромы нервы проникают через отверстия в боуменовой оболочке и образуют подэпителиальное сплетение [705, 971, 762, 763]. Иннервируются все эпителиальные клетки вплоть до поверхностных двух слоев, в которых нервные окончания имеют вид бусинок, колб Краузе, пластинок, лопаточек и др. [28, 29, 496, 762, 763, 878]. Концевые колбы Краузе, обеспечивающие температурную чувствительность, обнаруживаются лишь в области лимба. Некоторые сплетения нервных волокон контактируют с клетками Ларгенганса [971, 762, 763]. Иннервации дес-
Роговая оболочка и склера

|
|
| / КГ Г V |
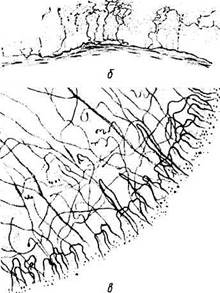
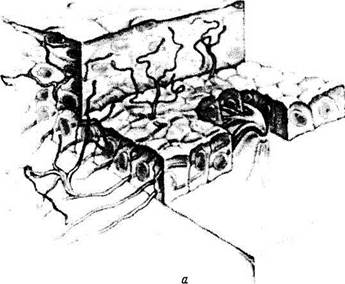
Рис. 3.2.12. Схематические изображения особенностей иннервации роговой оболочки:
а — трехмерное изображение прохождения и распределения нервных волокон в роговой оболочке; б—поперечный срез роговицы. Распределение нервных волокон и нервных окончаний в переднем эпителии роговой оболочки; в — плоскостной препарат. Поверхностное краевое нервное сплетение
цеметовой оболочки и эндотелия не выявляются [931].
Ультраструктурные особенности нервов роговой оболочки позволяют некоторым авторам предполагать наличие пептидэргической иннервации как кератоцитов, так и эпителиальных клеток [762, 763].
Время регенерации нервных волокон роговицы длится около трех месяцев. Начинается регенерация нервов с периферии роговицы по направлению к центру. Помимо чувствительной иннервации, роговица обеспечена и вегетативной. Вегетативные волокна исходят из трех ганглиев. Это тройничный, ресничный и верхний шейный ганглии. Основным доказательством наличия вегетативной иннервации роговой оболочки является обнаружение отхождения нервных веточек от нервов лимбальных сосудов [1, 2, 28], а также эспериментальные исследования по перерезке нервных стволов, отходящих от вегетативных узлов, или после «раздражения» последних. Вегетативная иннервация обеспечивает трофику роговицы. Денервация роговой оболочки в эксперименте путем перерезки нервных стволов, входящих в глазное яблоко вблизи зрительного нерва, приводит к развитию дистрофических процессов, напоминающих нейропаралитический кератит у человека [16,17, 30]. Аналогичного характера дистрофические процессы роговой оболочки и структур переднего отдела глаза наблюдаются и после проведения циркляжа силиконовой лентой, которая передавливает ресничные нервы [12].
О значении иннервации говорит и то, что одним из необходимых основных условий диф-
ференциации эпителиальных и стромальных компонентов роговицы после травмы или кератопластики является реиннервация роговой оболочки [6, 30].
Старение роговой оболочки.Старение является естественным процессом у многоклеточных животных, приводящим к нарушению структуры и функции тканей и органов [923]. У человека признаки старения проявляются как функция времени. Выражаются они в нарушении дифференциации клеток, а их причиной являются биологические изменения, заложенные генетически или возникающие под влиянием на организм внешней среды.
Процесс старения ткани можно разделить на старение длительно существующих белков, старение делящихся клеток и старение неделя-щихся клеток [175].
Делящиеся клетки характеризуются тем, что их популяция поддерживается равновесием двух разнонаправленных процессов — скоростью размножения клеток и скоростью их гибели. Наиболее типичным примером такой популяции являются клетки переднего эпителия роговой оболочки. В роговичном эпителии полная смена дифференцированных эпителиальных клеток происходит за 5—7 дней [647, 695, 698]. Некоторые типы клеток способны к интенсивной пролиферации только после воздействия на них каких-либо внешних факторов. К подобным клеткам можно отнести кератоциты стромы роговицы [1115]. Клетки эндотелия роговицы также способны к делению, но деление эндотели-альных клеток у человека происходит исключительно редко [744, 977, 1087, 1127]. К неде-
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
лящимся клеткам относятся нейроны головного мозга, а в глазном яблоке нейроны сетчатой оболочки.
Старение роговицы приводит к возникновению различных структурных и функциональных изменений. Эти изменения приводят к нарушению прозрачности роговицы, снижению регенераторной способности, нарушению кривизны поверхности, снижению адаптационной способности ткани роговицы и др. [402].
Поскольку трудно отличить возрастные изменения от изменений, возникающих при различных патологических состояниях роговицы, имеет смысл кратко остановиться на основных положениях процесса старения.
Первоначально мы охарактеризуем особенности старения клетки.
Деление в популяции клеток происходит постоянно и находится под генетическим контролем [647, 997]. Одним из основных признаков старения клетки являются нарушение цикла репликации и нарушение жизненного цикла клетки. При этом клетки выходят из митотического цикла все в большем количестве. В клетках, вышедших из клеточного цикла, отмечается постоянное накопление нарушений как структуры, так и функции. Этим объясняется увеличение вероятности развития дегенерации ткани при старении по мере накопления подобных клеток [187, 455].
Необходимо указать на отличия между стареющими клетками и клетками, находящимися в состоянии покоя (О0-фаза). В состоянии покоя дифференцированные клетки не пролифе-рируют благодаря наличию контактного торможения. Стареющие клетки выходят из цикла не в состоянии конечной дифференциации [794]. Именно по этой причине фенотип дифференцированной и стареющей клетки, выходящей из митотического цикла, существенно отличается. В первую очередь необходимо указать на то, что стареющая клетка покидает цикл с содержанием ДНК, характерным фазе G! [395]. При этом в ее ДНК происходит ряд изменений, приводящих к нарушению функции клетки [455, 1004]. К основному нарушению относят подавление транскрипции части генов [1178]. Подобные изменения могут быть «критическими», т. е. изменениями, приводящими к нарушению целостности и функции всей ткани.
В литературе рассмаривается два основных пути, по которым происходит старение клетки. Первый путь — «конститутивное старение». Теория «конститутивного старения» предполагает, что при старении в результате пролиферации клеток увеличивается вероятность накопления в геноме ошибок, выводящих клетку из пролиферативного пула [837, 1015]. Этот процесс является вероятностным, и трудно определить закономерности его развития. Кинетику «конститутивного старения» можно объяснить возможным прогрессивно нараста-
ющим нарушением репликативной способности ДНК [55].
Второй путь старения клетки — это так называемое «реактивное старение». При этом типе старения предполагают, что, подобно апоп-тозу, старение может быть вызвано мутацией или влиянием на геном различных мутагенных факторов (противоопухолевые препараты и др.). Основным отличием от «конститутивного старения» является то, что подвергаются старению клетки с небольшой пролиферативной активностью. Офтальмолог должен помнить о подобном типе старения, поскольку в арсенале лечебных средств, используемых им, есть многочисленные мутагенные препараты, такие как 5-фтороурацил (применяется для предотвращения рубцевания после удаления птеригиума или после операции по поводу глаукомы), мито-цин С. Экспериментально показано ускорение процесса старения клеток под воздействием этих препаратов в культуре ткани [142]. Подобные лекарственные средства легко проникают через роговую оболочку и склеру при введении их в конъюнктивальную полость и могут явиться причиной преждевременного старения клеток различных структур глаза, что проявляется разным образом и спустя неодинаковый период времени после проведенного лечения [604, 570].
Представляют особый интерес и данные, указывающие на стимуляцию процессов старения кератоцитов стромы роговицы после удаления переднего эпителия. Выражается это резким увеличением явлений апоптоза кератоцитов стромы, особенно ее передних слоев. В последующем, после эпителизации роговицы, происходит замещение погибших клеток новыми кератоцитами, мигрирующими из задних слоев стромы [1173, 1174]. Гибель кератоцитов в такой ситуации является примером конститутивного старения. Подобный тип старения, сопровождающийся уменьшением плотности кератоцитов, может стать причиной развития хронических заболеваний роговой облочки различной этиологии.
К сожалению, явления старения структур роговой оболочки у человека изучены далеко не полностью. Тем не менее увеличение количества стареющих клеток с возрастом показано на культуре ткани клеток переднего эпителия, а также при исследовании роговой оболочки пожилых людей [374, 958]. С возрастом увеличивается также и количество старых клеток в эндотелии роговой оболочки [506].
Возникает вопрос: каким образом накопление с возрастом стареющих клеток влияет на частоту патологических состояний роговицы?
Основным изменением стареющей роговицы является снижение ее адаптационных возможностей. При этом роговица более подвержена инфекционному поражению. Увеличивается проницаемость как переднего, так и заднего эпителия [188].
Роговая оболочка и склера

 Нарушение распределения в эпителии роговицы интегринов приводит к нарушению межклеточных контактов, что является причиной более свободного проникновения в нее бактерий, вирусов и клеток крови [471, 489]. Выявлено также, что при старении нарушение целостности переднего эпителия роговицы сопровождается нарушением целостности и эндотелия [512]. Это, в свою очередь, приводит к отеку стромы роговицы и ее помутнению.
Нарушение распределения в эпителии роговицы интегринов приводит к нарушению межклеточных контактов, что является причиной более свободного проникновения в нее бактерий, вирусов и клеток крови [471, 489]. Выявлено также, что при старении нарушение целостности переднего эпителия роговицы сопровождается нарушением целостности и эндотелия [512]. Это, в свою очередь, приводит к отеку стромы роговицы и ее помутнению.
Исследований, посвященных изучению особенностей старения кератоцитов, немного. Тем не менее большинство исследователей переносят на эти клетки закономерности, выявленные при исследовании фибробластов in vitro. Показано, что при старении происходит экспрессия в фибробластах таких ферментов, как колагеназа, стромолизин и эластаза [398, 1223]. Наблюдается экспрессия металлопротеиназ [175, 740], уменьшение количества коллагена — mRNA [741]. Нарушен также синтез фибронек-тина [1023]; снижается синтез протеогликанов [512], а также способность фибробластов контролировать трехмерную организацию коллаге-новых волокон в культуре ткани. Отмечено накопление липофусцина в стареющих роговицах (cornea farlnata).
Дата добавления: 2015-03-26; просмотров: 1503;