Нервные окончания
Афферентные (чувствительные)нервные окончания (рис. 1.5.5, 1.5.6).
Афферентные нервные окончания представляют собой концевые аппараты дендритов чувствительных нейронов, повсеместно располагающихся во всех органах человека и дающие информацию центральной нервной системе об их состоянии. Воспринимают они раздражения, исходящие и из внешней среды, преобразуя их в нервный импульс. Механизм возникновения нервного импульса характеризуется уже описанными явлениями поляризации и деполяризации цитоплазматической мембраны отростка нервной клетки.
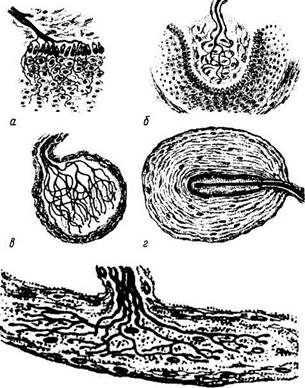
Существует ряд классификаций афферентных окончаний — в зависимости от специфичности раздражения (хеморецепторы, бароре-
Глава 1. КЛЕТКА И ТКАНИ

|
це Мейснера, терморецепторы колбы Краузе, тельца Руффини и др.).
Разнообразно строение рецепторов мышечной ткани, часть которых обнаруживается в наружных мышцах глаза. В этой связи на них мы остановимся более подробно. Наиболее распространенным рецептором мышечной ткани является нервно-мышечное веретено (рис. 1.5.6). Это образование регистрирует растяжение волокон поперечно-полосатых мышц. Представляют они собой сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено располагается вдоль мышечных волокон. Веретено покрыто тонкой соединительнотканной капсулой (продолжение периневрия), внутри которой находятся тонкие поперечнополосатые интрафу-зальные мышечные волокна двух видов:
— волокна с ядерной сумкой — в расши
ренной центральной части которых содержатся
скопления ядер (1—4 — волокна/веретено);
— волокна с ядерной цепочкой — более тон
кие с расположением ядер в виде цепочки в
центральной части (до 10 волокон/веретено).
Рис. 1.5.5. Особенности строения различных рецеп-торных окончаний:
а — свободные нервные окончания; б— тельце Мейснера; в — колба Краузе; г — тельце Фатер—Пачини; д — тельце Руффини
цепторы, механорецепторы, терморецепторы и др.), от особенностей строения (свободные нервные окончания и несвободные).
Обонятельные, вкусовые, зрительные и слуховые рецепторы, а также рецепторы, воспринимающие движение частей тела относительно направления силы тяжести, называют специальными органами чувств. В последующих главах этой книги мы подробно остановимся только на зрительных рецепторах.
Рецепторы разнообразны по форме, строению и функциям. В данном разделе нашей задачей не является подробное описание различных рецепторов. Упомянем лишь о некоторых из них в разрезе описания основных принципов строения. При этом необходимо указать на различия свободных и несвободных нервных окончаний. Первые характеризуются тем, что они состоят только из ветвления осевых цилиндров нервного волокна и клетки глии. При этом они контактируют разветвлениями осевого цилиндра с клетками, возбуждающими их (рецепторы эпителиальных тканей). Несвободные нервные окончания отличаются тем, что в своем составе они содержат все компоненты нервного волокна. Если они покрыты соединительнотканной капсулой, они называются инкапсулированными (тельце Фатер—Пачини, осязательное тель-
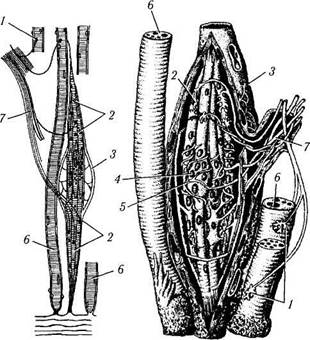
Рис. 1.5.6. Строение нервно-мышечного веретена:
а—моторная иннервация интрафузальных и экстрафузальных мышечных волокон; б — спиральные афферентные нервные окончания вокруг интрафузальных мышечных волокон в области ядерных сумок (/ — нервно-мышечные эффекторные окончания экстрафузальных мышечных волокон; 2 — моторные бляшки интрафузальных мышечных волокон; 3 — соединительнотканная капсула; 4 — ядерная сумка; 5 — чувствительные кольцеспираль-ные нервные окончания вокруг ядерных сумок; 6 — скелетные мышечные волокна; 7 — нерв)
Строение периферической нервной системы.
Чувствительные нервные волокна образуют кольцеспиральные окончания на центральной части интрафузальных волокон обоих типов и гроздьевидные окончания у краев волокон с ядерной цепочкой.
Двигательные нервные волокна — тонкие, образуют мелкие нервно-мышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.
Рецепторами растяжения мышцы являются также нервно-сухожильные веретена (сухожильные органы Гольджи). Это веретеновид-ные инкапсулированные структуры длиной около 0,5—1,0 мм. Располагаются они в области соединения волокон поперечнополосатых мышц с коллагеновыми волокнами сухожилий. Каждое веретено образовано капсулой из плоских фиброцитов (продолжение периневрия), которая охватывает группу сухожильных пучков, оплетенных многочисленными терминальными веточками нервных волокон, частично покрытых леммоцитами. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
Эфферентные нервные окончания несут информацию от центральной нервной системы к исполнительному органу. Это окончания нервных волокон на мышечных клетках, железах и др. Более подробное их описание будет приведено в соответствующих разделах. Здесь мы подробно остановимся лишь на нервно-мышечном синапсе (моторная бляшка). Моторная бляшка располагается на волокнах поперечнополосатых мышц. Состоит она из концевого ветвления аксона, образующего пресинапти-ческую часть, специализированного участка на мышечном волокне, соответствующего постси-наптической части, и разделяющей их синапти-ческой щели. В крупных мышцах один аксон иннервирует большое количество мышечных волокон, а в небольших мышцах (наружные мышцы глаза) каждое мышечное волокно или их небольшая группа иннервируется одним аксоном. Один мотонейрон в совокупности с ин-нервируемыми им мышечными волокнами образует двигательную единицу.
Пресинаптическая часть формируется следующим образом. Вблизи мышечного волокна аксон утрачивает миелиновую оболочку и дает несколько веточек, которые сверху покрыты уплощенными леммоцитами и базальной мембраной, переходящей с мышечного волокна. В терминалах аксона имеются митохондрии и синапти-ческие пузырьки, содержащие ацетилхолин.
Синаптическая щель имеет ширину 50 нм. Располагается она между плазмолеммой ветвлений аксона и мышечного волокна. Содержит она материал базальной мембраны и отростки глиальных клеток, разделяющих соседние активные зоны одного окончания.
Постсинаптическая часть представлена мембраной мышечного волокна (сарколеммой), об-
разующей многочисленные складки (вторичные синаптические щели). Эти складки увеличивают общую площадь щели и заполнены материалом, являющимся продолжением базальной мембраны. В области нервно-мышечного окончания мышечное волокно не имеет исчерченности, содержит многочисленные митохондрии, цистерны шероховатого эндоплазматического ретику-лума и скопление ядер.
Механизм передачи нервного импульса на мышечное волокно сходен с таковым в химическом межнейронном синапсе. При деполяризации пресинаптической мембраны происходит выделение ацетилхолина в синаптическую щель. Связывание ацетилхолина с холинорецеп-торами в постсинаптической мембране вызывает ее деполяризацию и последующее сокращение мышечного волокна. Медиатор отщепляется от рецептора и быстро разрушается ацетил-холинэстеразой.
Дата добавления: 2015-03-26; просмотров: 1218;