Нервная ткань
Основной задачей данного раздела является изложение сведений о клеточной и тканевой организации нервной системы.
В центральной нервной системе различают две группы клеточных элементов — нейроны и нейроглии.
Нейроны(рис. 1.4.28, 1.4.29) состоят из тела клеток и отростков. В зависимости от количества отростков различают псевдоуниполяр-ные (биполярные нейроны, у которых два отростка вблизи тела клетки приближаются друг к другу, в результате чего складывается впечатление о существовании только одного отростка), биполярные (нейроны, имеющие два отростка — аксон и дендрит) и мультиполярные нейроны (нейроны, от тела которых отходит более двух цитоплазматических отростков — аксон и многочисленные дендриты).
Независимо от количества отростков нейроны всегда содержат один отросток, по которому информация передается от нейрона к исполняющему органу. Этот отросток имеет различную длину и может достигать нескольких десятков сантиметров. Называется он аксоном (нейрит).
Остальные отростки, а их число может быть самым различным, отличаются тем, что по ним
Ткани

| :|Ш.;;.:. |
| * .V'.' V НИ •'•-• Y '- |

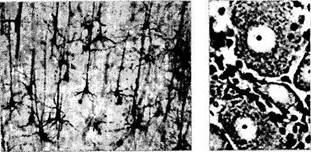

Рис. 1.4.28. Микроскопическое строениеразличных типов нейронов:
|
А — пирамидные клетки коры головного мозга; 6—нейроны ганглия (ганглиозные клетки). В цитоплазме нейронов определяется тигроидное вещество (вещество Ниссля). Окружаютнейроны клетки-сателлиты; в — корзинчатые клетки мозжечка; г — муль-типолярные нейроны коры головного мозга, в цитоплазме которых видны нейрофиламенты
'\
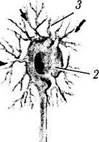
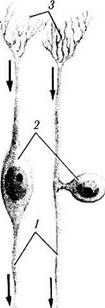
Рис. 1.4.29. Основные морфологические типы нейронов:
а — мультиполярный нейрон; б — биполярные нейроны (/ — аксон: 2 — тело клетки; 3 — дендриты)
информация подходит к нейрону. Эти отростки дихотомически ветвятся, в связи с чем получили название дендриты.
Нейроны довольно существенно отличаются по строению от клеток других тканей. Это, в первую очередь, относится к телу нейрона. Тела нейронов обычно крупные и могут иметь самую разнообразную форму. Ядро, как правило, располагается в центре клетки, содержит незначительное количество зерен гетерохрома-тина и обладает 2—3 крупными четкими круглыми ядрышками. Эти особенности отражают высокую активность процессов транскрипции в ядре нейрона. Около ядрышка в нейронах у женщин часто выявляется тельце Барра — крупная глыбка хроматина, содержащая конденсированную Х-хромосому.
Цитоплазма нейронов содержит большинство известных органоидов и окружена плазмо-леммой, способной к проведению нервного импульса вследствие локального тока Na+ в цитоплазму и К+ из нее через потенциал-зависимые мембранные ионные каналы.
Для нейронов характерны и специфические черты организации цитоплазмы. К таковым можно отнести наличие вещества Ниссля (тигроидное вещество, тигроид) (рис. 1.4.28, 1.4.29). Вещество Ниссля обнаруживается в крупных нейронах (ганглиозные клетки сетчатки) и представляет собой крупные базофильные внутри-цитоплазматические глыбки. Ультраструктур-но показано, что вещество Ниссля есть не что иное, как насыщение цитоплазмы цистернами гранулярного эндоплазматического ретикулума, содержащими обилие свободных и связанных рибосом и полирибосом. Это указывает на исключительно высокий белковый метаболизм в нейронах. Важно подчеркнуть, что при развитии патологических состояний, сопровождающихся повреждением отростков нейрона или тела, вещество Ниссля исчезает (хроматолиз). Нередко хроматолиз является первым признаком развивающегося заболевания.
Второй особенностью организации цитоплазмы нейрона является наличие нейротрубочек, нейрофиламентов (промежуточные филаменты) (рис. 1.4.30, 1.4.31).
Нейрофиламенты диаметром 10 нм складываются в пучки, выполняющие цитоплазму клетки.
Нейротрубочки выполняют опорную функцию, особенно в цитоплазматических отростках, и имеют типичное строение. Их диаметр 24 нм.
К другим особенностям строения нейрона необходимо отнести и возможность наличия в их цитоплазме двух типов пигмента — меланина (substantia nigra) и липофусцина.
Особенности строения отростков нервных клеток имеет смысл рассматривать в связи с выполняемыми ими функциями.
Как указывалось выше, аксон (нейрит) может иметь довольно большую длину (от 1 мм
Глава 1. КЛЕТКА И ТКАНИ

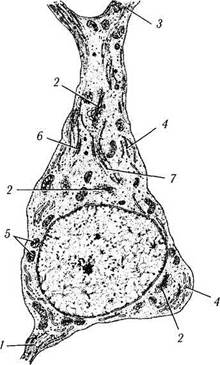
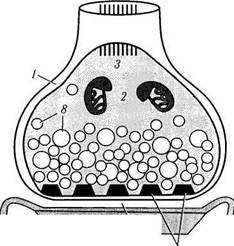
Рис. 1.4.30. Схема ультраструктурной организации нейрона (по А. Хэм, Д. Кормак, 1983):
1— аксонный холмик; 2— комплекс Гольджи; 3 — дендриты; 4 — гранулярный эндоплазматический ретикулум; 5 — митохондрии; 6—микротрубочки; 7 — филаменты

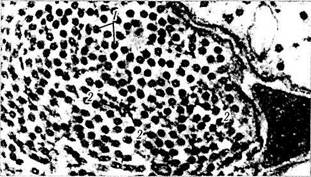
Рис. 1.4.31. Нейротрубочки и нейрофиламенты нейронов:
а — продольный срез слоя нервных волокон сетчатки (по Hogan et al., 1971); б — поперечный срез слоя нервных волокон сетчатки (/ — нейротрубочки; 2— нейрофиламенты)
до 1,5 ж у человека). Аксон отходит от утолщенного участка тела нейрона, не содержащего хромофильной субстанции, — аксонного холмика, в котором генерируются нервные импульсы. Он почти на всем протяжении покрыт гли-альной оболочкой. Аксон может по своему ходу давать ответвления (коллатерали), которые обычно отходят от него под прямым углом. В конечном участке аксон нередко распадается на тонкие веточки (телодендрии). Аксон заканчивается специализированными терминаля-ми (нервными окончаниями) на других нейронах или клетках рабочих органов.
Отличается по строению и цитоплазма аксона. Центральная часть цитоплазмы аксона (ак-соплазма) содержит большое количество аксо-плазматических пузырьков, нейрофиламентов, микротрубочек, ориентированных продольно. Гранулярный эндоплазматический ретикулум, аппарат Гольджи отсутствуют. Встречаются редкие рибосомы.
Таким образом, видно, что белковый метаболизм в аксоне определяется обменом, происходящим в теле клетки. Поскольку объем аксона может быть во много десятков раз больше объема тела нейрона, метаболизм в аксоне поддерживается специальным механизмом — ак-сонным транспортом.
Различают два типа аксонного транспорта — антероградный (из тела нейрона по аксону) и ретроградный (направлен от терминалей в сторону тела клетки). В свою очередь антероградный транспорт подразделяют на медленный (скорость потока по аксону 1—5 мм в сутки) и быстрый (5—10 мм в час).
Медленный поток несет пузырьки, лизосо-мы и ферменты, участвующие в синтезе нейро-медиаторов (норадреналина). Быстрый поток несет некоторые ферменты, участвующие в выполнении синаптической функции, гликопротеи-ды, фосфолипиды, митохондрии и нейросекре-торные гранулы.
Ретроградный аксонный транспорт (100— 200 мм/сут.) способствует удалению веществ из области терминалей, возвращению пузырьков, митохондрий.
Феномен транспорта используется для изучения межнейронных связей путем введения маркеров в область расположения терминалей или клеточных тел и выявления областей его последующего распространения описанными механизмами.
Дендриты, в отличие от аксона, дихотомически делятся. Крупные дендриты отличаются от аксона тем, что они содержат рибосомы и цистерны гранулярного эндоплазматического ретикулума, а также нейротрубочки, нейрофиламенты и митохондрии. Существует и дендритный транспорт со скоростью, примерно равной быстрому потоку в аксоне.
После описания структуры нейронов логично остановиться на механизмах передачи нерв-
Ткани

|
|
| \ ..■■■■ / ' ' V - ■ |
| / |
| 7 4 |
ного импульса. Связь нервных клеток между собой осуществляется с помощью синапсов.
Синапсы.Синапсы подразделяются на электрические и химические.
Синапсы могут различным образом располагаться на нейроне. В тех случаях, когда аксон оканчивается на дендрите, синапс называют аксодендритным, а когда на теле нейрона— аксосинаптическим. Аксоны, заканчивающиеся на других аксонах, называются аксо-аксональными (рис. 1.4.32).
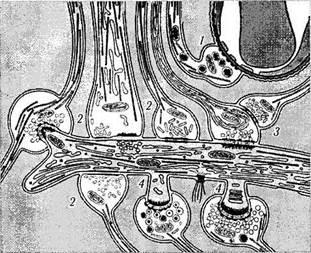
Рис. 1.4.32. Схематическое изображение различных типов синаптических соединений между нейронами:
/ — аксосоматический синапс; 2 — аксодендритные синапсы различных типов; 3 — аксоаксонный синапс; 4 — аксодендритные синапсы с дендритным шипиком
Электрические синапсы в центральной нервной системе млекопитающих редки. Довольно большое их количество в сетчатой оболочке. Эти синапсы имеют строение щелевых соединений, в которых мембраны синаптически связанных клеток (пре- и постсинаптическая) разделены промежутком шириной 2 нм, пронизанным коннексонами. Последние представляют собой трубочки, образованные белковыми молекулами и служащие водными каналами, через которые мелкие молекулы и ионы могут транспортироваться из одной клетки в другую. Когда потенциал действия, распространяющийся по мембране одной клетки, достигает области щелевого соединения, электрический ток пассивно протекает через щель от одной клетки к другой. Импульс способен передаваться в обоих направлениях и практически без задержки.
Химический синапс при ультраструктурном исследовании характеризуется следующим (рис. 1.4.33). Между параллельно расположенными пре- и постсинаптическими мембранами двух нейронов имеется пространство шириной 20—30 нм, называемое синаптической щелью. Пресинаптическая часть образуется аксоном по его ходу (приходящий синапс) или
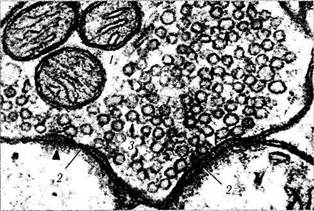
Рис. 1.4.33. Схема строения синапса (а) и его ультраструктурные особенности (б):
а — схема строения синапса (/ — пресинаптическая часть; 2— митохондрии; 3 — нейротрубочки и нейрофиламенты; 4 — пресинаптическая мембрана с пресинаптическими уплотнениями;
5 — постсинаптическая мембрана; 6 — постсинаптическое уплот
нение; 7 — синаптическая щель; 8 — синаптические пузырьки);
6 — ультраструктурные особенности синапса (/ — митохондрии;
2 — постсинаптическое уплотнение; 3 — синаптические пузырьки)
представляет собой расширенную конечную часть аксона (концевой бутон). В ней содержатся митохондрии, гладкая эндоплазматичес-кая сеть, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20—65 нм, в которых находится нейромедиатор. Нейро-медиаторы вырабатываются в теле клетки и механизмом быстрого транспорта переносятся в окончание аксона. На внутренней стороне плазмолеммы, обращенной к синаптической щели (пресинаптической мембраны), имеется пресинаптическое уплотнение, образованное фибриллярной гексагональной белковой сетью, ячейки которой способствуют равномерному распределению синаптических пузырьков по поверхности мембраны.
Глава 1. КЛЕТКА И ТКАНИ
Синаптическая щель выполнена жидкостью, содержащей гликопротеиды, гликолипиды, а также электронноплотные частицы. Иногда она содержит гликопротеиновые интрасинапти-ческие филаменты толщиной 5 нм, которые являются элементами специализированного гли-кокаликса, обеспечивающими адгезивные связи пре- и постсинаптической частей, а также направленную диффузию медиатора.
Постсинаптическая часть представлена постсинаптической мембраной, содержащей особые комплексы интегральных белков — синаптичес-кие рецепторы, связывающиеся с нейромедиа-тором. Мембрана утолщена за счет скопления под ней плотного филаментозного белкового материала (постсинаптическое утолщение).
Процесс передачи нервного импульса схематически представляется следующим образом. Поступление нервного импульса в окончание пресинаптического нейрона к синапсу проявляется в деполяризации пресинаптической мембраны, что делает мембрану проницаемой для ионов кальция. При этом ионы кальция начинают поступать в пресинаптическую цитоплазму. Это приводит к слиянию синаптических пузырьков, содержащих нейромедиатор, с пресинаптической мембраной. Слившиеся мембраны открываются в синаптическую щель, и содержимое пузырьков изливается туда. Медиатор, выделившийся в синаптическую щель, и воздействует на постсинаптическую мембрану, делая ее более проницаемой для ионов натрия, проникающих в цитоплазму постсинаптическо-го нейрона. При этом потенциал покоя уменьшается и происходит деполяризация постсинаптической мембраны и генерируется нервный импульс. Подобный тип передачи нервного импульса характерен возбудительным нейронам. В тормозных нейронах высвобождение медиатора в синаптическую щель приводит к открытию каналов для ионов хлора, которые и переходят в цитоплазму постсинаптического окончания. При этом увеличивается ее отрицательный потенциал (гиперполяризация), а возбудимость клетки понижается. Таким образом, суммарное действие обоих видов синапсов на один нейрон приводит к определенному балансу между двумя противоположными видами синаптических эффектов. Медиаторами, опосредующими возбуждение, например, служат ацетилхолин и глютамат, а торможение опосредуется ГАМК и глицином.
После прекращения взаимодействия медиатора с рецепторами постсинаптической мембраны большая часть его эндоцитозом захватывается пресинаптической частью, меньшая рассеивается в пространстве и захватывается глиальными клетками. Некоторые медиаторы расщепляются ферментами на компоненты, которые далее захватываются пресинаптической частью. Затем они повторно используются для образования новых синаптических пузырьков.
В заключение необходимо привести также сведения относительно биохимической классификации нейронов. Основана эта классификация на химическом составе нейромедиатора того или иного нейрона. Выделяют холинергичес-кие (медиатор — ацетилхолин), адренергические (медиатор — норадреналин), серотонинергичес-кие (медиатор — серотонин), дофаминергичес-кие (медиатор — дофамин), ГАМК-ергические (медиатор — гамма-аминомасляная кислота), пуринергические (медиатор — АТФ и его производные), пептидергические (медиаторы — субстанция Р, энкефалины, эндорфины, вазоактив-ный интерстинальный пептид, холецистокенин, нейротензин, бомбезин и другие нейропептиды) нейроны. В некоторых нейронах терминали содержат одновременно два типа нейромедиатора. Различные типы нейронов, отличающиеся химическим составом нейромедиатора, обнаруживаются в сетчатой оболочке, как и в центральной нервной системе (см. Сетчатка).
Вторую группу клеток нервной ткани составляют клетки нейроглии.
Нейроглия.Нейроглия — термин, обозначающий обширную группу клеток нервной ткани, обеспечивающих деятельность нейронов и выполняющих опорную, трофическую, разграничительную, барьерную, секреторную и защитную функции.
Морфологически различают три типа гли-альных клеток — олигодендроциты, астроциты и микроглиальные клетки (рис. 1.4.34—1.4.36).
Олигодендроглия состоит из мелких клеток (олигодендроцитов) с короткими немногочисленными отростками. В цитоплазме клеток об-
Рис. 1.4.34. Различные типы глиальных клеток нервной ткани (по В. Л. Быкову, 1999):
1 — эпендимоцит; 2 — таницит; 3 — хориоидный эпендимоцит; 4 — протоплазматический астроцит; 5 — волокнистый астроцит; 6 — пограничная глиальная мембрана мозга; 7—периваскуляр-ная пограничная мембрана; 8 — клетки-сателлиты; 9 — нейрон; 10 — леммоцит (шванновская клетка); // — олигодендроцит; 12 — микроглиальная клетка и ее трансформация в процессе фагоцитоза
Ткани

|
| *Дендриты нейрона |
С■%
| ; |
наруживается хорошо развитый синтетический аппарат. Высокое содержание лизосом, митохондрий и гранул гликогена. В зависимости от особенностей строения олигодендроциты разделяют на крупные светлые, мелкие темные и промежуточные. У взрослых обнаруживаются преимущественно мелкие темные клетки. Длительность существования светлой клетки невелика, и она постепенно вызревает, превращаясь в малый темный олигодендроцит.
Одной из наиболее важных функций олиго-дендроцитов является образование миелиновой оболочки вокруг нервных волокон в центральной нервной системе. Миелин, выполняя функцию изолятора, участвует в процессе передачи по волокну нервного импульса.
Процесс миелинизации сводится к тому, что олигодендроцит обертывает своей цитоплазмой определенный участок нервного волокна последовательными спиральными слоями (рис. 1.4.37—1.4.39). В результате этого нервное волокно оказывается окутанным целым пакетом двухслойных цитоплазматических мембран, содержащих миелин. В результате этого и наступает изоляция нервного волокна. В миелинизации периферических нервов участвует шванновская клетка (леммоцит) (см. ниже).
Рис. 1.4.35. Светооптические особенности глиальных клеток:
а — астроцит и его отношение к дендритам нейрона; б — олиго-дендроциты головного мозга
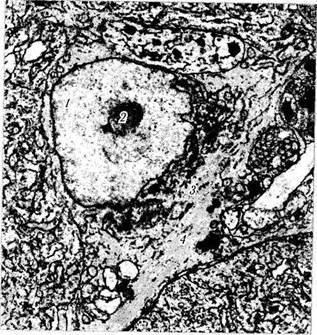
Рис. 1.4.36. Ультраструктурная организация глиальной клетки (олигодендроцит сетчатки):
/ ядро; 2 — ядрышко; 3 — цитоплазматические включения; 4—микрофибриллы
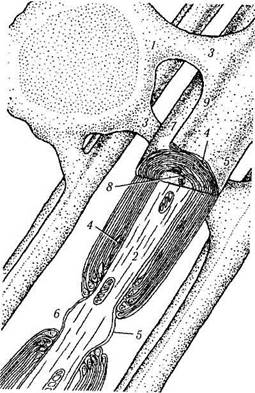
Рис. 1.4.37. Взаимоотношение олигодендроцита с нервным волокном и строение миелинизированного волокна (по Шаде, Форд, 1976):
1 — олигодендроцит; 2 — аксон; 3 — связь между телом клетки глии и миелиновой оболочкой; 4 — цитоплазма- 5 — цитоплазма-тическая мембрана; 6 — перехват Ранвье; 7 — петля плазматической мембраны; 8~ мезаксон; 9 — гребешок
Глава 1. КЛЕТКА И ТКАНИ


Рис. 1.4.38. Механизм формирования миелиновой оболочки шванновскими клетками (в периферической нервной системе) или олигодендроцитами (в центральной нервной системе)
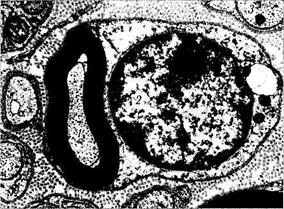
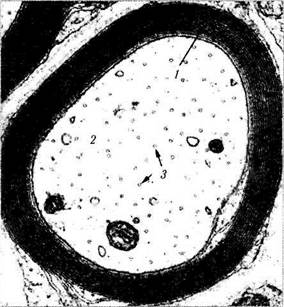
Рис. 1.4.39. Ультраструктурные особенности шваннов-ской клетки (а) и миелиновой оболочки периферического нерва (а, б):
а — отношение олигодендроцита к аксону (/—аксон; 2 — ядро
олигодендроцита; 3—немиелинизированный нервный ствол);
б—миелиновая оболочка аксона (/—миелиновая оболочка;
2— аксон; 3 — микротрубочки)
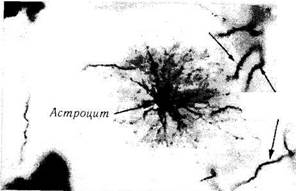
Астроглия представлена астроцитами (см. рис. 1.4.35, 1.4.36). Астроциты обладают многочисленными отростками, расходящимися от тела клетки в разных направлениях, напоминая при этом звезды.
Характеризуется клетка наличием светлого овального ядра. Ее цитоплазма содержит небольшое количество органоидов, но значительное количество зерен гликогена и промежуточных филаментов. Промежуточные филаменты содержат особый глиальный фибриллярный кислый белок (ГФКБ), который служит маркером астроцитов. Астроциты образуют щелевые соединения между собой, а также с клетками олигодендроглии и эпендимной глии.
Разделяют астроциты на волокнистые (фибриллярные) и протоплазматические (плазматические). Отличия между двумя типами клеток сводятся к тому, что цитоплаз-матические отростки фибриллярных астроцитов практически не ветвятся, в то время как у протоплазматических астроцитов ветвление хорошо выражено.
Ультраструктурная организация этих клеток приблизительно одинаковая. Пучки филаментов распространяются от одного отростка к другому, проходя через тело клетки, что создает ее особую жесткость.
Протоплазматические астроциты преимущественно встречаются в сером веществе центральной нервной системы, а волокнистые — в белом. Кроме того, волокнистые астроциты содержат большое количество ГФКБ.
Учитывая то, что клетки астроглии плотно контактируют с сосудами и нейронами, предполагают, что эти клетки выполняют опорную, разграничительную, транспортную, барьерную, трофическую, защитную и регулятор-ную функции.
Опорная функция сводится к формированию астроцитами каркаса, внутри которого располагаются нейроны и волокна. В ходе эмбрионального развития они служат опорными и направляющими элементами, вдоль которых происходит миграция нейронов. Направляющая функция связана также с секрецией ростовых факторов, распознаваемых нейронами.
Разграничительная, транспортная и барьерная функции астроглии сводятся к участию в образовании гемато-энцефалического и нейро-ликворного барьеров, на которых более подробно мы остановимся в 4-й главе.
Трофическая функция является наиболее важной функцией астроглии. Направлена она на поддержание определенных концентраций ионов кальция и медиаторов в окружении нейронов. Астроциты совместно с олигодендрогли-ей принимают участие в метаболизме медиаторов, активно захватывая их из синаптической щели и передавая нейрону.
Защитная функция астроглии сводится к участию в различных защитных реакциях — фагоцитозе, иммунной реакции, репаративной.
Как и олигодендроциты, астроциты способны к пополнению клеточной популяции на протяжении всей жизни путем митотических делений клеток-предшественников. Их высокая про-
Ткани
 лиферативная активность способствует также глиальному рубцеванию поврежденных нервных тканей.
лиферативная активность способствует также глиальному рубцеванию поврежденных нервных тканей.
Микроглия — это мелкие клетки, разбросанные в белом и сером веществе мозга. Они составляют всего 5% популяции глиальных элементов. Микроглия обнаруживается и в сетчатой оболочке. Предполагают, что схожие по происхождению и функции клетки лежат в стекловидном теле вблизи сетчатки.
Считается, что микроглиальные клетки происходят из моноцитов или периваскулярных макрофагов мозга (мезенхимное происхождение). Структурной особенностью микроглии является насыщенность цитоплазмы лизосомами. Вероятней всего микроглия определяет защитную функцию в нервной системе, относясь к клеткам системы иммунитета. При патологических состояниях микроглиальные клетки проявляют способность к передвижению, фагоцитозу. Их количество существенно увеличивается при воспалительных и дегенеративных заболеваниях нервной ткани. При этом они утрачивают отростки, округляются и способны фагоцитировать остатки погибших клеток. При повышении активности микроглиальных клеток усиливается секреция ряда цитокинов и токсических радикалов. Именно с этим связывают усиленную гибель нейронов путем апоптоза при некоторых заболеваниях нервной системы.
К глиальным элементам относят и эпендим-ную глию {эпендима). Клетки эпендимной глии выстилают желудочки мозга и спинномозговой канал. К эпендимной глие ряд авторов относит и плоские клетки, выстилающие мозговые оболочки (менинготелий).
Особенностью эпендимоцитов является наличие на апикальной поверхности ресничек, которые при своем движении перемещают спинномозговую жидкость. Клетки эпендимной глии плотно прикрепляются друг к другу межклеточными соединениями. Часть клеток лежит на базальной мембране. Некоторые клетки от ба-зальной своей поверхности отдают отростки по направлению мозга, входящие в состав поверхностной пограничной глиальной мембраны (краевая глия).
Эпендимная глия выполняет опорную, трофическую, барьерную и секреторную функции. Барьерная функция эпендимной глии сводится к участию в образовании нейро-ликворного и гемато-ликворного барьеров.
Покрывает эпендима и сосудистые сплетения мозга (специализированными клетками — хориоидными эпендимоцитами и таницитами).
Завершая описание строения структурных элементов центральной нервной системы, необходимо упомянуть и о нейропиле, т.е. структурном компоненте центральной нервной системы, представляющем собой при световой микроскопии светло-голубой материал, в который погружены нейроны, их отростки, капиллярные сосу-
ды. Ультраструктурно показано, что нейропиль представляет собой переплетение клеточных тел, отростков нейронов и глиальных элементов.
1.5. СТРОЕНИЕ ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
В периферической нервной системе различают следующие компоненты:
1. Ганглии.
2. Нервы.
3. Нервные окончания и специализирован
ные органы чувств.
Дата добавления: 2015-03-26; просмотров: 1161;