ХАРАКТЕРИСТИКА ВІРУСОПОДІБНИХ БІОПОЛІМЕРІВ ТА ВІРУСІВ 13 страница
Различия в пищевом поведении различных видов животных несомненно связаны с особенностями их вкусовой чувствительности.
Неоднократно высказывались предположения, что четыре качества вкусовых ощущений человека вызываются раздражением четырех видов рецепторов, представленных
227
в разнообразных комбинациях в каждом из вкусовых сосочков и с различной плотностью распределенных на отдельных участках языка. Хорошо известно, что у человека кончик языка более чувствителен к сладким веществам, а основание, наоборот, — к горьким. Ощущение соленого связано с раздражением боков и кончика языка, а ощущение кислого возникает, главным образом, при раздражении области, ограниченной боковыми сторонами языка. Можно было бы далее предположить, что вкусовая информация передается в центральную нервную систему по волокнам, каждое из которых объединяет рецепторы нескольких видов, и это объясняет возможность возникновения смешанных вкусовых ощущений. Однако в настоящее время отсутствуют прямые доказательства правильности этой точки зрения.
В порядке возрастания вкусовой чувствительности основные вкусовые ощущения располагаются следующим образом: соленое, кислое, сладкое и горькое. Ощущение горечи во рту возникает при концентрации хинина равной всего лишь 3•107 молей на литр дистиллированной воды. Для получения ощущений сладкого, соленого или кислого концентрация раствора должна быть по крайней мере в 1000 раз больше. Это обстоятельство имеет очевидный биологический смысл, так как горькие вещества часто бывают ядовитыми.
Дифференциальные пороги вкусовой чувствительности равны примерно 0,20. При этом, закону Бугера-Вебера подчиняется только ощущение горького.
При нанесении химических веществ на определенные участки языка было обнаружено, что вкусовые ощущения подвержены быстрой и полной адаптации. Для этого обычно бывает достаточно 30 секунд. Так чувствительность к соленому падает за это время в 300 раз, а к сладкому в 20 раз. Изменение порогов к кислому и горькому выражено менее сильно.
В естественных условиях адаптация не возникает, так как в результате движений языка происходит постоянное изменение химической стимуляции отдельных рецепторов. Таким образом,движения языка способствуют более точному распознаванию вкусовых веществ. В оценке вкуса пищи большую роль играют также обонятельные,
228
тактильные, температурные и болевые компоненты. На пороги вкусовых ощущений влияют факторы, вызывающие сенсибилизацию (см. стр. 83 и др.). Известно, что на вкусовую чувствительность влияет температура, причем, это влияние по-разному сказывается на каждом из четырех вкусовых качеств. При различных заболеваниях пищеварительного, дыхательного и других аппаратов, участвующих в поддержании постоянства внутренней среды организма, так или иначе поражается и вкусовая чувствительность.
О зависимости вкусовой чувствительности от равновесия обменных процессов в организме говорят данные, полученные при изучении выбора крысами солевых растворов. Оказалось, что обычно крысы предпочитают 40%-ный раствор глюкозы, смешанный с 1%-ным раствором поваренной соли, чистому 40%-ому раствору глюкозы. Однако, если в брюшную полость крыс вводили 10 мл физиологического раствора, содержащего около 1% поваренной соли, то животные предпочитали чистый раствор глюкозы. Таким образом, присутствие в организме избытка поваренной соли сразу уменьшает "солевой" аппетит животных.
Вкусовая чувствительность человека также заметно уменьшается при насыщении и возрастает при голодании.
Вкус и обоняние, как протопатические виды чувствительности, имеют более аффективную окраску чем все остальные ощущения, за исключением боли. При этом интенсивное ощущение горького неприятно, а сладкого — приятно.
Обонятельные ощущения
В отличие от вкусовой чувствительности спектр различных воспринимаемых человеком запахов исключительно разнообразен. Обычно, определяя тот или иной запах, мы стремимся уловить его сходство с другими, известными нам запахами, например, запахом сирени, моря или уксуса. Однако, по-видимому, язык слишком беден, чтобы описать воспринимаемые запахи. Поэтому классификация запахов представляет собой трудную, далеко нерешенную задачу.
Исследователи неоднократно пытались описать и систематизировать
229
запахи. Наиболее ранние попытки сводились к простому перечислению различных запахов. В середине восемнадцатого века шведский натуралист К. Линней предложил классификацию, включающую девять растительных запахов. Эту классификацию можно было использовать как подсобное средство при отнесении растений к тому или иному классу.
Одна из наиболее известных классификаций принадлежит немецкому психологу Х. Хеннингу (1924), подробно исследовавшему более четырехсот различных запахов. Х. Хеннинг пришел к выводу, что существует шесть основных запахов: фруктовый, цветочный, смолистый, пряный, гнилостный и горелый. Отношения между ними принято изображать в виде треугольной призмы запахов, в углах которой расположены основные запахи (рис. 96). Все воспринимающиеся запахи расположены где-либо на границах, но не внутри призмы запахов.
Рис. 96. Призма запахов Х. Хеннинга.
Однако последующие работы других исследователей показали, что хотя призма Х. Хеннинга передает общие отношения между запахами, она недостаточно точна. В частности, испытуемые часто называли в качестве основного запаха кислый, а некоторые запахи помещали внутри призмы. Поэтому была предложена другая схема, включающая четыре основных компонента: ароматный, кислый, горелый и гнилостный, интенсивность которых оценивается по условной
230
шкале от нуля до восьми. Эта классификация в настоящее время имеет известное практическое применение.
Запахи можно классифицировать и по другим основаниям. Ранее уже говорилось о том, что Э. М. Хорнбостелю удалось успешно разделить "светлые" и "темные" запахи (см. стр. 57). О тесной связи обонятельных ощущений с вкусом свидетельствует существование "сладких" и "кислых" запахов. Важное практическое значение имеет разделение приятных и неприятных запахов. Эта классификация используется в парфюмерном производстве.
Физическим условием пахучести вещества является его летучесть, так как только непосредственно контактирующие с рецепторной поверхностью молекулы вещества могут вызвать запах. Огромная роль обоняния в регуляции пищевого поведения объясняет тот факт, что за небольшим исключением химическим условием пахучести является органическое происхождение вещества.
Основным нерешенным вопросом в изучении обоняния является вопрос о связи химической структуры вещества с его запахом и характером воздействия на хеморецепторы. Несмотря на имеющиеся описания соотношений химического строения и воспринимаемого запаха вещества, заранее предсказать по формуле вещества его запах трудно. Оказалось, что соединения самой различной структуры могут иметь сходные запахи, а очень близкие соединения могут пахнуть совершенно по-разному.
Было предложено немало теорий обоняния, однако ни одна из них не получила окончательного подтверждения.
Американским исследователем Дж. Эймуром (1964) была выдвинута "стереохимическая теория" обоняния. Она основана на предположении, что организм может различать формы молекул раздражителя. Дж. Эймур подразделил пахнущие вещества на несколько первичных запахов, описав характерные для них формы молекул. В соответствии с этой теорией специфическое ощущение запаха возникает в том случае, когда вещества с определенной формой молекулы попадают в подходящее "гнездо" на поверхности рецептора. Было выделено семь первичных запахов: камфорный, эфирный, цветочный, мятный, мускатный и гнилостный. Такие запахи, как запах миндаля, лимона, чеснока или запах "тухлого" являются
231
сложными, состоящими из двух или более первичных запахов.
В пользу этой теории говорит тот факт, что многие изомеры, т. е. вещества с одинаковым составом, но с разной формой молекулы, пахнут различно. С другой стороны, стереохимическая теория не объясняет, почему незначительные изменения концентрации вещества могут резко менять его запах.
В основе другой так называемой "вибрационной теории" обоняния лежит предположение о связи вибрационных движений молекулы пахнущего вещества и рецептора. В обонятельной мембране рецепторов был обнаружен пигмент желтого или коричневого цвета. Американский ученый Р. Райт (1964) высказал гипотезу, что молекулы пахнущего вещества, вибрируя, изменяют энергетический уровень пигмента. Однако и эта теория встретила много возражений.
Психофизические исследования показали, что, несмотря на затруднительный доступ к обонятельным рецепторам, присутствие пахнущего вещества может быть обнаружено даже при очень низких концентрациях. Для большинства запахов пороговая концентрация составляет от 10—7 до 10—14 молей на литр воздуха. Таким образом, из всех видов хеморецепции обоняние обладает наибольшей остротой. Оно в тысячи раз чувствительнее вкуса.
Дифференциальный порог обонятельной чувствительности колеблется в зависимости от вида запаха в пределах от 0,15 до 1,0. Это несколько больше, чем у других видов чувствительности, иными словами, замечаются лишь сравнительно большие изменения раздражителя.
На интенсивность запаха влияют температура и влажность, поскольку с ними связана летучесть вещества. Регулярные колебания обонятельной чувствительности могут достигать размаха 1 : 50. В начале дня и вечером люди более чувствительны к запахам, чем днем. Состояние насыщенности представляет собой фактор, заметно снижающий обонятельную чувствительность.
Чувствительность к запахам возрастает во время беременности, под действием некоторых фармакологических веществ, а также при раздражении обонятельного мозга.
Существуют тонкие индивидуальные различия в обонятельной
232
чувствительности здоровых людей. Например, некоторые люди не воспринимают ряд известных запахов. Это явление в известной мере аналогично явлению "цветовой" слепоты (см. стр. 123 и д.). Оно служит доводом в пользу предположения о существовании специализированных "детекторов" запахов.
Как хорошо известно, адаптация к запахам протекает очень быстро. Необходимым условием длительного сохранения обонятельных ощущений являются дыхательные движения, вызывающие циркуляцию воздуха в носовой полости. Эта функция дыхательных движений аналогична функции движения глаз в случае зрения и движений языка в случае вкуса.
При одновременном действии на обонятельные рецепторы молекул двух или нескольких пахучих веществ возможны эффекты маскировки, компенсации или слияния запахов. Маскировказаключается в том, что один из запахов, как правило, менее интенсивный, вообще не замечается испытуемым. Другая форма взаимодействия запахов — компенсация — состоит в том, что одновременное предъявление двух запахов может приводить к их взаимному ослаблению, так что никакого обонятельного ощущения не возникает. Наконец, сочетание двух или нескольких запахов может привести к возникновению качественно нового запаха, отличного от всех составляющих смесь пахучих веществ. На подобном слиянии основан эстетический эффект смешения запахов в парфюмерии — "букет запахов".
Было бы неверно считать, что обоняние человека определяется исключительно отологической потребностью в пище. Как уже отмечалось, при оценке запахов человек прежде всего сравнивает его с запахом знакомых ему предметов. Запахи для нас являются признаками огромного числа предметов и явлений. В природе насчитывается несколько десятков тысяч пахучих веществ. Комбинации их запахов могут быть бесконечно многообразными. Однако человек способен научиться различать самые тонкие запахи, коль скоро это требуется для выполнения его предметной, практической деятельности. Именно предметный характер обонятельных ощущений имел в виду Ф. Энгельс, писавший, что "собака, обладая значительно более тонким обонянием, чем человек, не различает
233
и сотой доли тех запахов, которые для человека являются определенными признаками различных веществ".х)
234
ВЕСТИБУЛЯРНАЯ СИСТЕМА
В филогенезе вестибуляная система появляется очень рано. Ее наиболее древняя часть, обеспечивающая получение информации о направлении силы гравитации, имеется в развитом виде у большинства беспозвоночных.
У рыб вестибулярная система дополняется аппаратом полукружных каналов, что позволяет ей принимать участие в регуляции движений организма. Свою роль важнейшего источника пространственной информации вестибулярная система сохраняет и у человека.
1. Анатомо-физиологическое строение вестибулярного аппарата
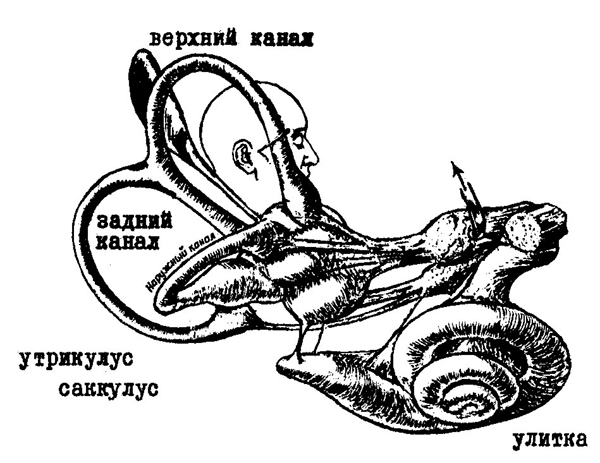
Рис. 97. Вестибулярный аппарат
(по С. Оксу, 1969).
С обеих сторон головы несколько спереди от ушной раковины в глубине костной ткани расположено внутреннее ухо — система ходов и полостей, называемая также из-за сложности своего строения лабиринтом. Внутреннее ухо функционально и анатомически
235
делится на две части: улитку, представляющую собой периферическое звено слуховой системы (см. стр. 177), и вестибулярный аппарат (рис. 97).
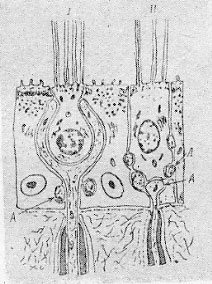
| Рис. 98. Два типа волосковых клеток (по С. Оксу, 1969) Видны окончания аксонов сенсорного нерва, подходящие непосредственно к телу рецептора (А). |
Вестибулярный аппарат состоит из двух отделов: полукружных каналов и статолитового органа. В них расположены высокоспециализированные механорецепторы — волосковые клетки. Свое название эти рецепторы получили благодаря выступающим на 0,03—0,04 мм над их наружной поверхностью волоскам, напоминающим реснички (рис. 98). Непосредственно к основанию волосковых клеток подходят окончания чувствительного нерва. Волосковые клетки вестибулярного аппарата выделяются среди других рецепторов двумя характерными свойствами. Во-первых, они обладают постоянной спонтанной активностью. Это позволяет кодировать сенсорную информацию не только с помощью увеличения частоты разрядов, но и за счет ее уменьшения. Во-вторых, волосковые клетки почти не подвержены адаптации, что исключительно важно для сохранения стабильности поступающих в ЦНС сведений о положении и движениях организма. Как показали электрофизиологические исследования
236
адекватным раздражителем волосковых клеток является касательная к их поверхности (тангенциональная) составляющая действующих на волоски деформирующих сил.
Сигналы о вращательных движениях головы поступают от аппарата полукружных каналов. Они представляют собой три расположенных во взаимноперпендикулярных плоскостях протока. Заполняющая полкружные каналы жидкость — эндолимфа — может перемещаться по ним в обоих направлениях. Быстрый поворот головы вызывает круговое движение эндолимфы в противоположную сторону. При этом эндолимфа увлекает находящийся в расширенной части каждого канала сгусток желеобразного вещества, называемый капулой. Смещения капулы вызывают деформацию чувствительных волосков, что приводит в зависимости от направления смещения к увеличению или к уменьшению частоты спонтанной импульсации. Эти изменения и несут мозгу информацию о движениях головы.
Благодаря расположению полукружных каналов в трех перпендикулярных плоскостях вращение головы в любом направлении однозначно кодируется картиной изменения импульсации от волосковых клеток различных каналов. С физической точки зрения подобная система имеет существенный недостаток, заключающийся в том, что она реагирует только на ускоренные движения головы. Поэтому продолжительное равномерное вращение испытуемого вокруг собственной оси должно привести к возвращению капул в "нулевое" положение, как при полном покое, а внезапная остановка — к резкому отклонению эндолимфы в направлении предшествовавшего движения, как при быстром рывке головы в противоположную сторону.
Однако обычно этот недостаток не мешает работе вестибулярной системы, так как практически всем естественным движениям головы свойственны частые смены направлений, большие, но кратковременные ускорения и отсутствие сколь-нибудь продолжительных периодов равномерного вращения. Кроме того, гидродинамические свойства эндолимфы таковы, что еще до раздражения волосковых клеток в вестибулярном аппарате осуществляется очень важное преобразование информации о движениях головы: благодаря своей вязкости и трению о стенки каналов эндолимфа играет роль механического интегратора ускорений головы во времени. В результате, отклонения капул, а следовательно и изменения активности рецепторов оказываются пропорциональными угловой скорости движения
237
головы (Н. Бишоф, 1966).
Другой отдел вестибулярного аппарата — статолитовый, или отолитовый орган — получил свое название из-за лежащих здесь на поверхности волосковых клеток известковых отолитов(статолитов). Если рецепторы полукружных каналов реагируют на угловую скорость движений головы, то отолитовый орган чувствителен к действующим на организм механическим силам. Эти силы бывают трех видов. Во-первых, на организм в обычных земных условиях постоянно действует сила гравитации. Для отражения направления этой силы, задающего вертикальное направление внешнего пространства, первоначально и возникла вестибулярная система. Затем, это инерционные и центробежные силы, возникающие при изменении скорости или направления движения головы.Результирующая всех этих сил является раздражителем отолитового органа. Передача столь сложной информации оказывается возможной благодаря специфическому строению отолитового органа.
Он состоит из двух частей: утрикулюса и саккулюса. Отолиты расположены в них под прямым углом друг к другу. У стоящего человека отолит утрикулюса лишь немного наклонен назад (на 10—20°) и занимает приблизительно горизонтальное положение, в саккулюсе же отолит вертикален. Под действием перечисленных выше сил отолиты деформируют волоски рецепторов, при этом сигналы от утрикулюса и саккулюса не повторяют, а дополняют друг друга.
Сенсорные импульсы от вестибулярного аппарата поступают в целый ряд инстанций головного мозга: мозжечок, средний мозг, таламус и кору. Столь широкие связи делают возможным использование вестибулярной информации для регуляции двигательных актов, например, таких, как сохранение равновесия, а также учитывать ее при перцептивной ориентации в пространстве. Особую роль, как известно (см. стр. 157 и д.), в создании образа интермодального внешнего пространства играют теменно-затылочные зоны коры. Не случайно они же являются кортикальными зонами представительства вестибулярного анализатора.
Моторные реакции на вестибулярную информацию
Вестибулярная система играет важную роль в координации движений субъекта. Возникающие в ответ на вестибулярную стимуляцию
238
моторные реакции выполняют две различные функции. Во-первых, они служат для сохранения нужной позы или движения тела вопреки сбивающим воздействиям со стороны внешних механических сил. Эти реакции называются реакциями сохранения равновесия. Во-вторых, вестибулярные рефлексы способствуют поддержанию неподвижности других органов чувств, прежде всего глаз, относительно координат внешнего пространства. Эти реакции называются компенсаторными.
Полукружные каналы и отолитовый орган, связанные с переработкой различной информации, отличаются также и по видам "запускаемых" движений. Раздражение полукружных каналов, как установил в конце прошлого века немецкий физиолог И. Эвальд, ведет к возникновению моторных тенденций движения в плоскости стимулируемого канала и в направлении движения эндолимфы. Эти эффекты представляют собой сохраняющие позу реакции в ответ на навязываемые извне нежелательные повороты, наклоны и колебания головы или всего тела. Несмотря на свой автоматический, рефлекторный характер эти реакции могут полностью отсутствовать, когда та же самая стимуляция полукружных каналов является следствием активно осуществляемых субъектом движений (Э. Ф. Хольст, 1950).
Влияние раздражения полукружных каналов на движения глаз выступает в форме вестибулярного нистагма. Как говорилось ранее (см. стр. 106), нистагмом называются движения глаз, состоящие из чередующихся саккадических (быстрая фаза) и следящих (медленная фаза) движений. Направлением нистагма обычно считают направление его быстрой фазы.
Запуск и регуляция этого типа движений осуществляется, главным образом, с помощью двух перцептивных систем: зрительной и вестибулярной. В обоих случаях функция нистагма состоит в том, чтобы максимально уменьшить смещения проекций окружающих предметов по сетчатке. Решающую роль при этом играет медленная фаза, которая всегда совпадает по направлению с относительным движением окружения. У животных с малоподвижными глазами (рептилии, птицы) нистагм протекает в форме характерных подергиваний головы при ходьбе.
При вестибулярном нистагме полукружные каналы регулируют
239
исключительно медленную фазу. С нее начинается нистагм и она определяет область, в которой он осуществляется (рис. 32, б). Быстрая фаза служит для возвращения глаза в исходную позицию. В естественных условиях вестибулярный нистагм носит далеко не столь регулярный характер: быстрая фаза практически отсутствует и наблюдаются лишь зеркальные по отношению к движениям головы гладкие компенсаторные движения.
Статолитовый орган, как уже отмечалось (см. стр. 237 и др.), в общем случае реагирует на результирующую трех различных сил: гравитационной, инерционной и центробежной. Подобный принцип работы биологически оправдан, т. к. именно эта результирующая определяет величину и направление фактически действующего на движущееся тело силового поля и является, таким образом, адекватным раздражителем по отношению к реакциям сохранения равновесия. Так велосипедист при повороте наклоняется в ту же сторону, ориентируя тело по результирующей центробежной и гравитационной сил, а не только по одной последней.
Существует несколько возможных путей сохранения равновесия тела. Все они входят в число регулируемых отолитовым аппаратом реакций. Во-первых, как в только что приведенном примере, это движения, изменяющие положение центра массы тела таким образом, чтобы его проекция не выходила за границы площади опоры. Вторая возможность заключается в изменении самой площади опоры, что мы делаем, когда, пытаясь сохранить равновесие, хватаемся за окружающие предметы. Наконец, третья возможность состоит в таком перераспределении тонуса мышечных звеньев, при котором тенденция к изменению относительного положения частей тела в данном силовом поле была бы наименьшей.
Разумеется, эти реакции возникают не только при участии вестибулярной системы. Особую роль играет при этом проприоцепция от мышц шеи, т. к. она позволяет дополнить вестибулярную информацию о положении головы информацией об относительном положении тела. Не случайно поэтому изменения мышечного тонуса наблюдаются не только при вестибулярной стимуляции, но и при увеличении
240
испульсации от проприоцепторов мышц шеи (например, за счет ее одностороннего нагревания: Р. Магнус, 1924).
Типичной компенсаторной реакцией, контролируемой отолитовым аппаратом, являются противовращения глаз во время боковых наклонов головы (рис. 99, б). Эта реакция служит для выпрямления положения проекции окружения на сетчатке. Максимальный поворот глаз достигает 5÷10°. Он наблюдается при наклоне головы, равном 60°. По отношению к отолитовым компенсаторным реакциям центробежные и инерционные силы представляют собой неадекватный биологический раздражитель. Поэтому противовращение глаз возникает и при вертикальном положении головы, когда испытуемый движется с ускорением в боковом направлении (рис. 99, в).
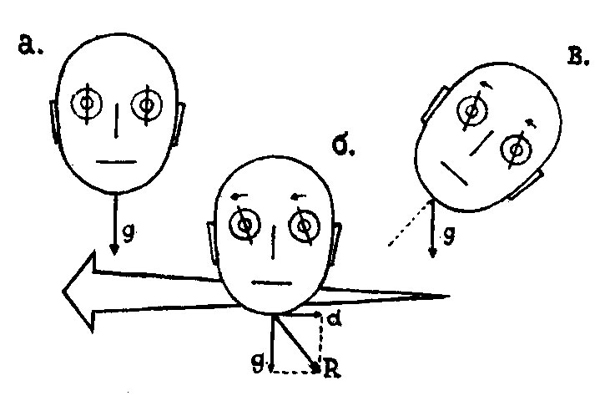
| Рис. 99. а) Нормальное положение глаз при вертикальной ориентации головы) — направление силы тяжести). б) Противовращение глаз при наклоне головы. в) Та же реакция, что и в случае (б), при ускоренном движении головы в направлении, указанном стрелой (d — инерционная сила). |
Восприятие положения и движения
Было бы ошибкой думать, что вестибулярная система участвует только в регуляции движений. Она является вторым по значению (после зрительной системы) источником пространственной информации, которая входит в качестве важной составляющей во все наши восприятия.
241
Повороты головы переменной скорости отражаются с помощью системы полукружных каналов. При этом любое смещение капулы ведет к восприятию собственного вращения в стабильном окружении. В естественных условиях наблюдается хорошее соответствие воспринятой таким образом и действительной угловой скоростью вращения головы.
Ту же самую информацию дает, как известно, и зрительная система. Кроме того, будет ли при данном раздражении вестибулярного аппарата воспринято движение всего тела или только головы определяется конкретным кинестетическим контекстом, в частности, проприоцептивной информацией от мышц шеи. Участие нескольких перцептивных систем в получении одной и той же информации влечет за собой два следствия. Во-первых, увеличивается надежность или избыточность восприятия пространственных характеристик. Во-вторых, становятся возможными интермодальные конфликты. Так, например, во время иллюзии индуцированного движения (см. стр. 161 и стр. 217) есть зрительная информация о движении тела, но нет вестибулярной. Во время качки на море или болтанки в самолете, напротив, есть вестибулярная информация о движении, но, как правило, нет полноценной зрительной. Проявлением таких конфликтов является головокружение и тошнота, т. е. хорошо известные симптомы "морской болезни".
Относительно простые взаимоотношения между движениями головы и их вестибулярным восприятием нарушаются когда испытуемый движется неестественным образом (см. стр. 139). Так внезапная остановка после продолжительного вращения вызывает у неподвижного испытуемого, сидящего на стуле с закрытыми глазами, отчетливое впечатление вращения в противоположную сторону. Одновременно появляется так называемый послевращательный нистагм, медленная фаза которого, как обычно, противоположна по направлению предполагаемому движению головы.
Вестибулярная система участвует также в восприятии движения объектов. Вестибулярная стимуляция вызывает видимое движение объекта, даже если он стабилизирован относительно сетчатки (Бифорд, 1966). Этот пример вновь доказывает, что системой отсчета для воспринимаемого движения служат реконструируемые на основании доступной организму информации координаты внешнего стабильного пространства.
242
Отолитовая система участвует в выполнении двух перцептивных действий: оценки положения тела и восприятия вертикального направления. В последние два десятилетия эта функция вестибулярной системы стала интенсивно исследоваться во многих странах мира. Проблема заключается в том, что оценивая вертикальное направление, совпадающее с направлением силы тяжести, отолитовый орган одновременно реагирует и на инерционные силы. Поэтому при ускоренном движении (в поезде, самолете, космическом корабле и т. д.) могут возникнуть различные вестибулярные иллюзии.
Так, например, при ускоренных движениях в горизонтальной плоскости в зависимости от направления ускорения у испытуемого создается впечатление наклонов вперед или назад (рис. 100). Эта иллюзия возникает потому, что за вертикаль, по отношению к которой оценивается положение тела, принимается результирующая сил тяжести и инерции.
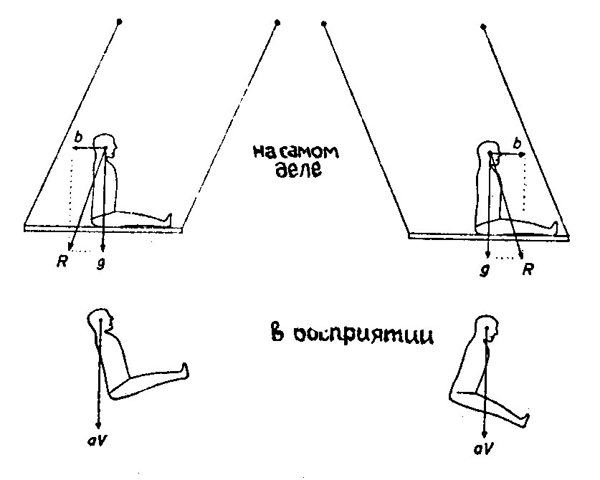
| Рис. 100. Восприятие положения при движении на параллельных качелях (по Л. Б. Джонкису, 1953) g — сила тяжести, b — инерционная сила, R —результирующая (раздражитель отолитового аппарата), αV — воспринимаемая вертикаль. |
243
Другая иллюзия, неоднократно отмечавшаяся космонавтами, по-видимому, связана с ориентацией статолитов в утрикулюсе. Поскольку они расположены несколько под углом к горизонтали (см. стр. 237), ускоренные движения в вертикальной плоскости (перегрузка или невесомость) приводят к возникновению инерционной силы, сдвигающей статолиты относительно волосковых клеток. Эта информация воспринимается человеком как опрокидывание на спину при перегрузке и наклон вперед при невесомости.
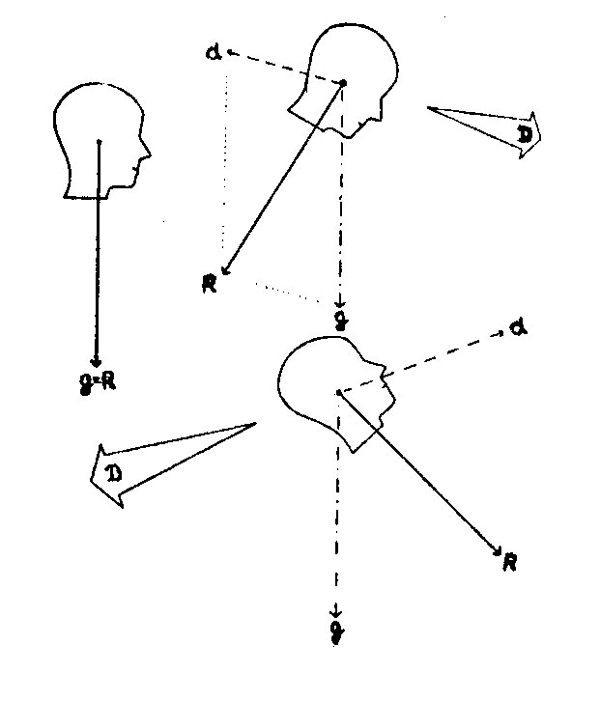
| Рис. 101. Неоднозначность вестибулярной информации о прямолинейном движении. На рисунке показаны три произвольно выбранные ситуации с индентичной стимуляцией отолитов. D — направление прямолинейного движения, d — инерционная сила, g — сила тяжести, R — результирующая. |
Одинаковая чувствительность и инерционным силам делает крайне ненадежной вестибулярную информацию, полученную в условиях полета на самолете. Например, на рис. 101 показан ряд ситуаций, в которых раздражение
244
вестибулярного аппарата будет одинаковым, несмотря на различное положение головы.
Исследование вестибулярной оценки вертикальности линий обычно проводилось в следующих условиях. В полной темноте испытуемому предъявлялась наклонная светящаяся линия, которую он должен был перевести в вертикальное положение. Оказалось, что испытуемый способен довольно успешно решать эту задачу даже, когда с помощью специального кресла его наклоняли в сторону. В связи с этим говорят о константности восприятия вертикали. Незначительные ошибки заключаются в том, что при малых углах собственного наклона испытуемый несколько переоценивает такой же наклон линии ("феномен Мюллера"), а при больших углах наклона (около 150°) — недооценивает его ("феномен Ауберта").
Недавно немецкие ученые Н. Бишоф и Х. Шерер (1971) провели серьезное экспериментальное исследование факторов, влияющих на оценку вертикальности линий в естественных условиях. Им удалось показать, что восприятия вертикальности обеспечивается совместной работой зрительной и вестибулярной систем. При этом зрительным признаком вертикального (и горизонтального) направления служит ориентация большинства контуров в зрительном поле. Действительно, доминирующие направления контуров обычно хорошо соответствуют горизонтали или вертикали (горизонт, деревья, дома и т. д.).
245
ХАРАКТЕРИСТИКА ВІРУСОПОДІБНИХ БІОПОЛІМЕРІВ ТА ВІРУСІВ
Дата добавления: 2015-03-19; просмотров: 963;