Симпатическая часть вегетативной нервной системы и мозговое вещество надпочечников
Катехоламины. Все три катехоламина естественного происхождения — норадреналин, адреналин и дофамин — функционируют как нейромедиаторы в центральной нервной системе. Норадреналин — нейромедиатор постганглионарных симпатических нервных окончаний, действует локально, в непосредственной близости от мест его высвобождения. Адреналин — циркулирующий в крови гормон мозгового вещества надпочечников — влияет на процессы, происходящие по всему организму. Существует также периферическая дофаминергическая система, но она не охарактеризована детально.
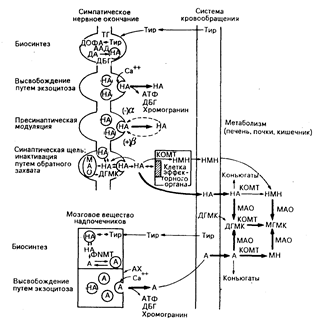
Рис. 66-1. Биосинтез, высвобождение и метаболизм катехоламинов. В верхней части рисунка схематически изображено периферическое симптоматическое нервное окончание; области луковицеобразных телец на терминальном волокне представлены расширениями, которые выявляют методом гистохимической флюоресценции как области высокой концентрации нейромедиаторов. В нижней части рисунка показаны хромаффинные клетки мозгового вещества надпочечников. Процессы биосинтеза, высвобождения, модуляции и обратного захвата показаны в последовательном порядке только в целях демонстрации: в организме все эти процессы протекают одновременно. ТГ — тирозингидроксилаза; ААД — декарбоксилаза ароматической L-аминокислоты;
ДА — дофамин; ДБГ — дофамин-р-гидроксилаза; НА — норадреналин; ФММТ — фенилэтаноламин-М-метилтрансфераза; А — адреналин; КОМТ — катехол-О-метилтрансфераза; НМН — норметанефрин; МАО— моноаминоксидаза; ДГМК — 3,4-дигидроксиманделиковая кислота; МГМК—З-метокси-4-гидроксиманделиковая кислота; АХ—ацетилхолин; Тир — тирозин; МН — метанефрин.
Биосинтез катехоламинов (рис. 66-1). Катехоламины синтезируются из аминокислоты тирозина, который последовательным гидрооксилированием преобразуется в диоксифенилаланин (дофа), затем декарбоксилируется, превращаясь в дофамин и гидроксилируется в положении боковой цепи, образуя норадреналин. Начальный этап этого процесса —- гидроксилирование тирозина — представляет собой этап, ограничивающий скорость всего процесса; он регулируется так, чтобы синтез диоксифенилаланина был связан с высвобождением норадреналина. Такая регуляция достигается изменениями как активности, так и количества тирозингидроксилазы. В мозговом веществе надпочечников и в тех центральных нейронах, в которых адреналин используется в качестве нейромедиатора, норадреналин N-метилируется в адреналин ферментом фенилэтаноламин-N-метилтрансферазой (ФNMT). Основная часть крови, снабжающей мозговое вещество надпочечников, обогащена кортикостероидами из коры надпочечников, и, поскольку надпочечниковая ФNМТ индуцируется глюкокортикоидами, способность мозгового вещества надпочечников образовывать адреналин может быть связана с ее стратегическим расположением внутри коры надпочечников.
Метаболизм катехоламинов (см. рис. 66-1). Основные метаболические трансформации катехоламинов включают в себя O-метилирование в метагидроксильной группе и окислительное дезаминирование. Процесс O-метилирования катализируется ферментом катехол-O-метилтрансферазой (КОМТ), а окислительному дезаминированию способствует моноаминоксидаза (МАО). КОМТ в печени и почках играет важную роль в метаболизме циркулирующих в крови катехоламинов. МАО, представляющая собой митохондриальный фермент, присутствующий в большинстве тканей, включая нервные окончания, менее значима в метаболизме циркулирующих в крови катехоламинов, но является важным фактором в регуляции их запасов в окончаниях периферических симпатических нервов. Метанефрины и 3-метокси-4-гидроксиманделиковая кислота (МГМК) являются главными конечными продуктами метаболизма и адреналина и норадреналина. Гомованилиновая кислота (ГВК) является конечным продуктом метаболизма дофамина.
Запасы катехоламинов и их высвобождение. Как в мозговом веществе надпочечников, так и в симпатических нервных окончаниях катехоламины сохраняются в субцеллюлярных гранулах и высвобождаются путем экзоцитоза. Большие запасы катехоламинов в этих тканях обеспечивают важный физиологический резерв, который поддерживает адекватное снабжение катехоламинами во время интенсивной стимуляции.
Мозговое вещество надпочечников. Хромаффинная ткань мозгового вещества обоих надпочечников здорового человека имеет массу около 1 г и содержит приблизительно 6 мг катехоламинов, 85% из которых приходится на долю адреналина. Высокая концентрация катехоламинов в хромаффинных гранулах поддерживается активным процессов их поступления, в котором участвуют мембраны гранул, и внутригранулярным комплексом хранения, в функционировании которого, по-видимому, участвуют АТФ, кальций и специфический белок гранул (хромогранин А). Секреция катехоламинов, стимулируемая ацетилхолином, высвобождающимся из преганглионарных симпатических нервов, возникает после попадания внутрь гранулы кальция, стимулирующего растворение мембраны хромаффинной гранулы и мембраны клетки; облитерация клеточной мембраны в месте ее растворения и вытеснение всего растворенного содержимого гранулы во внеклеточное пространство завершают процесс экзоцитоза (см. рис. 66-1). Ежесуточно происходит обмен приблизительно 2—10% общего запаса катехоламинов, содержащихся в мозговом веществе надпочечников.
Периферические симпатические нервные окончания. Периферические симпатические нервные окончания образуют сеть или сплетение, которое обеспечивает тесный контакт терминальных волокон с клетками, эффекторного органа. Весь норадреналин периферических тканей находится в симпатических нервных окончаниях, и густо иннервированные ткани содержат по 1— 2 мкг его на 1 г ткани. Норадреналин в нервных окончаниях находится в дискретных субцеллюлярных частицах, подобных хромаффинным гранулам мозгового вещества надпочечников. МАО, содержащаяся в миотохондриях нервных окончаний, играет важную роль в регуляции локальной концентрации норадреналина. Амины, находящиеся в луковицеобразных тельцах, защищены от окислительного дезаминирования; однако амины, локализованные внутри цитоплазмы, дезаминируются и превращаются в неактивные метаболиты. Высвобождение медиаторов из нервных окончаний происходит в ответ на раздражение и возникающие постсинаптические потенциалы передаются в виде импульсов по терминальным симпатическим волокнам (см. рис. 66-1).
Периферический адренергический нейроэффекторный синапс. Поглощение нейронами. Периферические симпатические нервные окончания участвуют в системе транспорта аминов, активно поглощая их из внеклеточной жидкости. Субстратами для этого процесса служит ряд синтетических аминов и аминов естественного происхождения. Поглощением нейронами, или обратным захватом, высвобожденного локально норадреналина заканчивается действие этого нейромедиатора и поддерживается постоянство его запасов (см. рис. 66-1).
Пресинаптическая модуляция. Характер взаимосвязи между прохождением импульсов по нервному волокну и высвобождением норадреналина находится под влиянием целого ряда факторов. Так, пониженная температура и ацидоз снижают количество норадреналина, высвобождаемого в ответ на импульсы симпатического нерва. Некоторые химические медиаторы воздействуют на периферическое симпатическое нервное окончание (определяемое как место пресинаптического действия), модифицируя характер передачи импульсов по симпатическим нервным волокнам путем влияния на количество норадреналина, выделяемого в ответ на нервные импульсы. Пресинаптическая модуляция может быть как подавляющей, так и усиливающей. Определенные модуляторы, такие как катехоламины и ацетилхолин, могут или угнетать, или усиливать высвобождение норадреналина, вызывая антагонистические эффекты, опосредуемые соответственно через различные адренергические или холинергические рецепторы. К веществам, оказывающим угнетающее действие на высвобождение норадреналина в пресинаптическом нервном .окончании, относятся: катехоламины (2-адренорецептор), ацетилхолин (мускариновый рецептор), дофамин (D2-рецептор), гистамин (Н2-рецептор), серотонин, аденозин, энкефалины и простагландины. К числу усиливающих пресинаптических модуляторов относятся: катехоламины (бета2-адренорецептор), ацетилхолин (никотиновый рецептор) и ангиотензин II. Общее значение пресинаптической модуляции, так же как и относительную степень важности различных медиаторов, еще предстоит выяснить.
Пресинаптические адренорецепторы. Катехоламины уменьшают высвобождение норадреналина, действуя через Пресинаптические -адренорецепторы по классической системе негативной обратной связи. Регуляция посредством обратной связи осложняется тем, что активация адренорецепторов усиливает высвобождение норадреналина. Были выдвинуты две гипотезы физиологического интегрирования антагонистических воздействий - и адренорецепторов на высвобождение норадреналина. Одна из них заключается в том, что адренорецепторы опосредуют реакции, развивающиеся при более низких концентрациях агониста, чем реакция, опосредуемые через -адренорецепторы, в то время как -адреноопоссредуемые реакции доминируют при более высоких концентрациях агониста. Поэтому при низких уровнях симпатической стимуляции, когда концентрации норадреналина в синаптической щели низки, адреноопосредуемая положительная обратная связь может доминировать, в результате чего его высвобождение усилится. И наоборот, при высоких уровнях симпатической стимуляции; когда концентрация норадреналина в симпатической щели высока, доминирует -адреноопосредуемая обратная связь и происходит угнетение высвобождения норадреналина. Другая гипотеза состоит в том, что Пресинаптические адренорецепторы более чувствительны к адреналину, чем к норадреналину; поэтому уровни содержания циркулирующего в крови адреналина могут стимулировать Пресинаптические адренорецепторы, тем самым увеличивая высвобождение норадреналина и усиливая симпатическую нейропередачу.
Пресинаптические холинергические рецепторы. Несмотря на то что было описано как угнетающее, так и усиливающее действие ацетилхолина на высвобождение норадреналина, угнетающее действие первого, опосредуемое через мускариновый холинергический рецептор, происходит при более низких его концентрациях и, вероятно, имеет большое физиологическое значение. Это периферическое угнетающее действие ацетилхолина на адренергическую нейропередачу может усиливать реципрокные изменения центральных парасимпатических и симпатических эфферентных импульсов, возникающих при регуляции различных физиологических реакций.
Дата добавления: 2015-03-17; просмотров: 1291;