И.Т КРУГЛИКОВА
СТРОЕНИЕ КОЖИ
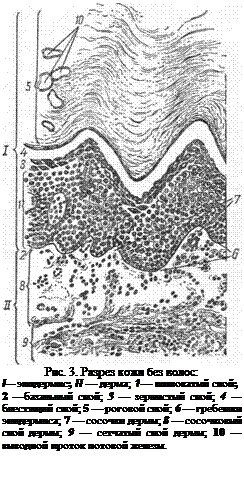
Кожа — corium — млекопитающих в основном состоит: из надкожницы, или эпидермиса, и основы кожи, или дермы. Там, где кожа легко смещается и собирается в складки, имеется подкожный слой, при помощи которого кожа связывается с нижележащими органами (рис. 3).
 Эпидермис — epidermis — наружный слой кожи (/). У млекопитающих эпидермис образован многослойным плоским эпителием. В местах, где кожа не покрыта волосами,
Эпидермис — epidermis — наружный слой кожи (/). У млекопитающих эпидермис образован многослойным плоским эпителием. В местах, где кожа не покрыта волосами,
эпидермис обычно составляет около 1—2% по отношению ко всей толщине кожи. В нем можно различить несколько слоев. Наиболее глубокими слоями являются базальный и шиповатый. Базальный слой представлен одним слоем высоких, призматических, энергично размножающихся клеток, среди которых встречаются
содержащие пигмент меланоциты, обусловливающие окраску кожи. В местах соприкосновения с нижележащими слоями базальные клетки несут микроворсинки, благодаря которым улучшаются условия обмена веществ. Шиповатый слой слагается из нескольких пластов (около 10) клеток,
которые образуются из более старых призматических клеток, потерявших связь с базальной мембраной и вытесненных сюда благодаря давлению молодых, энергично растущих базальных клеток. Шиповатые клетки размножаются менее активно, соприкасаются друг с другом при помощи протоплазматических отростков, вместе их встречи лежат десмосомы, разделенные пространством, заполненным тканевой жидкостью. Через нее происходит обмен веществ, а благодаря присутствию вязких мукополисахаридов тканевая жидкость прочно связывает клетки друг с другом. В цитоплазме шиповатых клеток имеются тонофибриллы, которые заканчиваются в десмосомах; они придают упругость эпидермису.
Зернистый слой (3) образован 2—4 слоями клеток, которые образовались из шиповатых. Это живые клетки, в их цитоплазме имеются крупные гранулы кератогиалина, хорошо красящиеся ядерными красителями. Зернистые клетки смещаются к поверхности кожи, формируя следующий блестящий слой ее.
Блестящий слой (4) слагается из 2—3 пластов уже мертвых уплощенных клеток, лишенных ядер. Их цитоплазма вытеснена элеидином, образовавшимся из кератогиалина зернистых клеток. Границ клеток в данном слое не видно. Изменяясь, эти клетки постепенно смещаются в самый наружный роговой слой кожи.
Роговой слой (5) самый толстый и состоит из нескольких десятков слоев совершенно мертвых клеток, приобретающих вид роговых чешуек. В каждой чешуйке есть оболочка, состоящая из настоящего рогового вещества — кератина и полости, заполненной воздухом или жироподобной массой. В силу такого строения чешуйки обладают упругостью и малой теплопроводностью. Поверхностные чешуйки рогового слоя теряют связь с глубоколежащими клетками эпидермиса и отторгаются от кожи, заменяясь вновь возникающими клетками. Задержка отторгаемых клеток в волосяном
покрове сельскохозяйственных животных ведет к нарушению многообразной функции кожи. Поэтому необходимо кожный покров животных систематически чистить и мыть.
Там, где кожа покрыта волосами, эпидермис значительно тоньше и состоит только из базального, шиповатого и рогового слоев (рис. 4).
Основа кожи, или собственно кожа — dermis, cutis, s. corium — развивается из дерматома мезодермы и состоит из соединительной ткани. В ней различают сосочковый и сетчатый слои (рис. 3—II—8, 9).
Сосочковый слой расположен под базальной мембраной, отделяющей основу кожи от эпидермиса. На поверхности его имеются сосочки (7), лежащие между гребешками эпидермиса. Этот слой состоит из ретикулярной и рыхлой соединительной ткани. В нем находятся отдельные пучки гладкой мышечной ткани и пигментные клетки, определяющие цвет кожи. В коже, несущей волосы, сосочковый слой основы кожи или совсем не образует сосочков, или они очень незначительны, и тогда граница между
эпидермисом и основой кожи представляется в виде прямой или слегка волнистой линии. Сосочковый слой без резкой границы переходит в сетчатый.
Сетчатый слой (9) состоит из плотной соединительной ткани. Клеток в нем мало, зато много эластических волокон и мощных коллагеновых пучков, переплетающихся между собой в виде плотной сетки. Дерма придает коже прочность. Именно из этого слоя изготовляют различные кожевенные изделия.
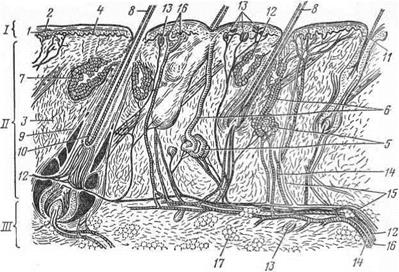 Рис. 4. Разрез кожи с волосами:
I —эпидермис; 1 — базальный и 2 — роговой слои; II — дерма; 3—сетчатый слой дермы; 4—сосочки дермы; 5—потовые железы; 6 — их выводные протоки; 7—сальные железы; 8 — стержень волоса; 9—волосяная сумка; 10—волосяные влагалища; 11 — сменяющийся волос; III—подкожный слой; 12—нервы; 13—кожные рецепторы; 14 — артерии; 15 — вены; 16 — лимфатические сосуды; 17 — жировая ткань.
Рис. 4. Разрез кожи с волосами:
I —эпидермис; 1 — базальный и 2 — роговой слои; II — дерма; 3—сетчатый слой дермы; 4—сосочки дермы; 5—потовые железы; 6 — их выводные протоки; 7—сальные железы; 8 — стержень волоса; 9—волосяная сумка; 10—волосяные влагалища; 11 — сменяющийся волос; III—подкожный слой; 12—нервы; 13—кожные рецепторы; 14 — артерии; 15 — вены; 16 — лимфатические сосуды; 17 — жировая ткань.
|
Подкожная основа (клетчатка) —tela subcutanea —(рис. 4—III)состоит из рыхлой соединительной или ретикулярной ткани и содержит большое количество жировых долек (17). Благодаря этому подкожный слой является важнейшим жировым депо, смягчает механические воздействия и обеспечивает возможность смещения кожи. У полусальных свиней жировой слой в области 6—7-го грудного позвонка достигает толщины 4—5 см, у сальных свыше 7 см и называется шпиком. У курдючных овец жир откладывается на ягодицах и у корня хвоста, формируя курдюк. У верблюда отложения жира образуют горб.
Хорошо развитый подкожный слой обусловливает большую подвижность кожи. Отсутствие или малая прослойка подкожного слоя, наоборот, ведет к плотному соединению основы кожи с подлежащими частями тела животного, к отсутствию или очень слабой подвижности кожи.
Сосуды кожи в сетчатом и сосочковом слоях образуют сплетения, причем ветвями последнего в каждом сосочке формируется густая капиллярная сеть, которая диффузно осуществляет питание эпидермиса. Между артериями и венами кожи имеются анастомозы, участвующие в терморегуляции.
Лимфатические сосуды особенно развиты в подкожном слое.
Нервы кожи представлены большим количеством чувствительных нервных окончаний и волокон. Последние образуют густое сплетение в сосочковом слое дермы. Нервные окончания многочисленны и неравномерно распределены. Благодаря им кожа в организме играет роль органа осязания.
Кожа образует ряд производных. Главные из них волосы, сальные и потовые железы, а также молочная железа, пальцевые мякиши, копыто, рога.
РАЗВИТИЕ И СТРОЕНИЕ ВОЛОСА
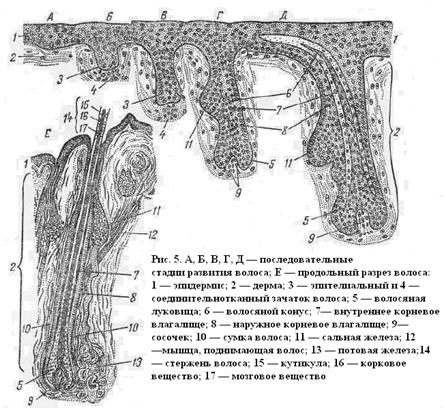 Развитие и рост волоса (pilus) начинается с утолщения эпителия, затем благодаря усиленному размножению клеток образуется зачаток, который в виде клеточного тяжа, все увеличивающегося в размере, погружается в
Развитие и рост волоса (pilus) начинается с утолщения эпителия, затем благодаря усиленному размножению клеток образуется зачаток, который в виде клеточного тяжа, все увеличивающегося в размере, погружается в
дерму кожи (рис. 5). На конце клеточного тяжа образуется утолщение,
формирующее луковицу (5). Внутрь луковицы врастает мезенхима, образующая сосочек волоса (9). Из средней части эпителиального зачатка образуется выпячивание, дающее начало сальной железе. В области луковицы клетки усиленно размножаются и дают начало стержню волоса, который, раздвигая клетки эпителиального тяжа, пробивает себе путь к поверхности
кожи.
Под давлением молодых клеток, непрерывно образующихся в луковице, стержень продолжает удлиняться. Клетки эпителия, расположенные около внутрикожной части стержня, формируют наружное (8) и внутреннее(7) корневые влагалища. Из окружающей мезенхимы, кроме волосяного сосочка, вокруг корня волоса развивается соединительнотканная сумка волоса (10), а также гладкая мускулатура (12).
Строение волоса. Сложившийся волос состоит из стержня и корня.
Стержень (14) имеет надкожную часть (шерсть, или «волос») и часть, скрытую в коже, входящую в состав корня волоса. На поперечном разрезе стержня видно, что он состоит из мозгового и коркового вещества, а также кутикулы (15). Сердцевина, или мозговое вещество (17), занимает центральное место в стержне. Это сплошной или прерывистый продольный тяж, состоящий из одного или нескольких рядов многоугольных (полигональных) живых, частично ороговевших клеток эпителиального происхождения. В цитоплазме клеток отлагается кератогиалин, который ближе к наружному слою стержня превращается в элеидин, а последний в кератин. Внутри клеток и между ними обнаруживают пузырьки воздуха, поэтому волос обладает небольшой теплопроводностью, защищая тело животного от охлаждения. В сердцевине находится пигмент, придающий цвет волосу. Самые нежные волосы овец — пух, волосы молодых животных и некоторые другие не имеют мозгового вещества. Наоборот, у оленя оно развито очень сильно. Корковое вещество (16) — основная масса стержня, придает ему механическую прочность, растяжимость, гибкость и упругость. Волосы, лишенные
этого слоя (северный олень), легко рвутся. В корневой части, около луковицы, стержень состоит из живых клеток с округлыми ядрами и слабыми признаками ороговения. По мере удаления от луковицы клетки становятся удлиненными и более ороговевшими, причем роговое вещество образуется сразу в виде твердого кератина, обусловливающего механические свойства стержня. В надкожной части сильно вытянутые клетки коркового слоя обычно уже лишены ядер, но содержат пигмент. С возрастом в корковом веществе увеличивается количество пузырьков газа, из-за чего волос седеет. Кутикула волоса (15) состоит из плоских, ороговевших, налегающих одна на другую, подобно черепице, или тесно прилегающих друг к другу безъядерных клеток. От конфигурации границ клеток кутикулы зависит разнообразный рисунок волоса, неодинаковый у животных различных пород. Кутикула защищает волос от действия влаги, света, химических веществ
и механических повреждений. От ее свойств зависят прядильные качества шерсти.
Корень волоса состоит: из внутрикожной части стержня, окутывающего его волосяного фолликула, луковицы с волосяным сосочком, а также сальных желез и мышцы. Внутри кожи часть стержня вблизи луковицы состоит из живых, малоороговевших клеток. Ближе к поверхности кожи он построен так же, как и наружный стержень.
Волосяной фолликул окутывает внутрикожную часть стержня и, подобно коже, состоит из эпителия и соединительной ткани. Из эпителия образуются внутреннее и наружное корневые влагалища, а из соединительной ткани — волосяная сумка. Внутреннее корневое влагалище (7) прилегает к стержню. Оно начинается с луковицы и доходит обычно лишь до места впадения сальных желез. У коров оно достигает поверхности кожи и является продолжением рогового слоя ее. Данное влагалище слагается из нескольких слоев клеток. Самый внутренний слой — кутикула влагалища,
состоящая из одного ряда ороговевших клеток, Следующий слой — гранулосодержащий эпителиальный, представлен 1—2 рядами светлых клеток с небольшим количеством трихогиалина. За этим слоем идет бледный эпителиальный слой, в котором при обычной окраске ядра различить невозможно. Наружное корневое влагалище (8) является прямым продолжением базального слоя эпидермиса кожи и имеется на всем протяжении корня волоса. Количество составляющих его клеток по направлению к луковице постепенно уменьшается. Самые наружные, более высокие клетки влагалища — производящие. Благодаря им образуются остальные клетки наружного влагалища.
Волосяная сумка (10) образована соединительной тканью с хорошо выраженными внутренними циркулярными и наружными продольными коллагеновыми и эластическими волокнами. Сумка хорошо развита в толстых волосах. От волосяной сумки к эпидермису кожи направляются под тупым углом пучки гладких мышечных клеток, которые, сокращаясь, поднимают волосы, а также содействуют выведению секрета сальных желез.
Луковица волоса (5) слагается из живых, усиленно размножающихся эпителиальных клеток. Они дают материал для образования стержня и внутреннего корневого влагалища.
Волосяной сосочек (9) состоит из соединительной ткани с массой кровеносных сосудов и нервов. Первые доставляют необходимые для жизнедеятельности волоса питательные вещества, а нервы обеспечивают связь волоса с организмом.
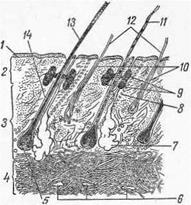
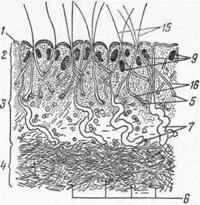
Корень с поверхностью кожи образует некоторый угол, поэтому стержень ложится более или менее параллельно поверхности кожи. Если волосяной мешок зигзагообразно изогнут, то выдвигающийся из него наружу стержень имеет большую или меньшую извитость. У тонкорунных овец из полости мешка может выходить несколько стержней (рис. 6—5), которые склеиваются секретом кожных желез в косички, а последние объединяются в штапель.
Периодически происходит смена волос. При этом производящие клетки луковицы начинают получать из сосочков меньше питательных веществ, они перестают делиться, часть их ороговевает и вместе со стержнем отделяется от живых клеток луковицы. Оказавшись свободным, стержень вместе с участком луковицы выдвигается из волосяного
 | |||
 | |||
мешка наружу и отпадает. Из остатка сосочка и живых клеток луковицы развивается новый волос, помещающийся в том же волосяном мешке.
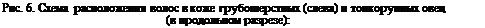 |
1—эпидермис; 2 — подэпидермальная зона сосочкового слоя; 3 — промежуточная зона сосочкового слоя;4— ретикулярный слой; 5 — луковица и сосочек; 6—кровеносный сосуд; 7—секреторные отделы потовых желез; 8 — мышца; 9 — дольки сальных желез; 10—вторичные фолликулы; 11—переходный волос;12 — пух; 13—ость; 14— первичный фолликул; 15—шерстяные волокна; 16 —фолликул.
СТРОЕНИЕ САЛЬНЫХ И ПОТОВЫХ ЖЕЛЕЗ
Сальные железы — glandulae sebaceae — располагаются преимущественно в основе кожи; по строению они относятся к простым, часто разветвленным альвеолярным железам (рис. 6 и 7—А, Б). Секреторные отделы их не имеют полостей. Наружные клетки их плоские или кубические, являются производящими. Энергично размножаясь, они постепенно оттесняются к центру альвеолы. По мере приближения к центру в них все
больше и больше накапливается вязкий жирный секрет. В результате ядро и часть цитоплазмы редуцируются, и такие переродившиеся клетки по протоку выделяются на поверхность кожи (кожное сало). Некоторые клетки стенки альвеолы уплощаются, ороговевают и в виде чешуек примешиваются к секрету.
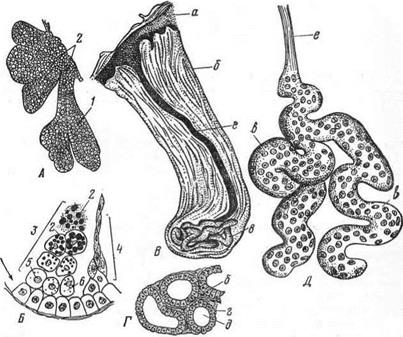 Рис. 7. Кожные железы млекопитающего:
Продольный разрез сальной железы при слабом (А) увеличении и образование жирового секрета в участке концевого отдела (Б); 1 — концевой отдел железы; 2 — секрет; 3 — последовательные стадии образования жирового секрета; 4 — образование роговых чешуек; 5 — ядро железистой клетки; 6 — цитоплазма ее; В — общий вид потовой железы; Г — концевой отдел (клубочек) потовой железы лошади (поперечный разрез); Д —общий вид потовой железы коровы; а — эпидермис кожи; б — соединительная ткань кожи; в — концевой отдел; г — эпителий концевого отдела; д — просвет концевого отдела; е — выводной проток железы.
Рис. 7. Кожные железы млекопитающего:
Продольный разрез сальной железы при слабом (А) увеличении и образование жирового секрета в участке концевого отдела (Б); 1 — концевой отдел железы; 2 — секрет; 3 — последовательные стадии образования жирового секрета; 4 — образование роговых чешуек; 5 — ядро железистой клетки; 6 — цитоплазма ее; В — общий вид потовой железы; Г — концевой отдел (клубочек) потовой железы лошади (поперечный разрез); Д —общий вид потовой железы коровы; а — эпидермис кожи; б — соединительная ткань кожи; в — концевой отдел; г — эпителий концевого отдела; д — просвет концевого отдела; е — выводной проток железы.
|
Таким образом, по типу секреции эти железы — типично голокриновые. Протокасальных желез в большинстве случаев открываются в волосяные фолликулы и изредка самостоятельными отверстиями на поверхности кожи (например, на головке полового члена самца). Протоки выстланы многослойным плоским эпителием. Сальных желез нет в соске вымени коровы, в коже носового зеркальца хищников и мелких жвачных, на мякишах, рогах, копытах и некоторых других местах тела животного.
Потовые железы — glandulae sudoriferae (рис.7—В,Д) — участвуют в терморегуляции и выведении продукта азотистого обмена — мочевины. Они лежат в глубоких слоях дермы, заходя частично в подкожный слой (рис.4—6). По своему строению относятся к простым, иногда ветвящимся, трубчатым железам, по типу секреции — к мерокриновым, а железы отдельных участков тела (например, паховой области) — к апокриновым железам. Секреторный отдел железы у коровы слабо изогнут, у лошади и свиньи он не ветвится, но очень длинен и свернут в клубочек
(рис. 7—В, Г, Д). Стенки этого отдела имеют два слоя клеток: в наружном находятся миоэпителиальные клетки, а внутренний составляют кубические железистые клетки. Выводные протоки желез слегка извитые и также имеют два слоя клеток. Открываются они в большинстве случаев в волосяные фолликулы, реже — возле волосяных воронок, а в некоторых безволосых местах — прямо на поверхности кожи потовыми порами.
Кроме сальных и потовых желез, в некоторых местах кожного покрова сельскохозяйственных животных лежат другие железы. Так, у крупного рогатого скота в носо-губном зеркальце имеются серозные железы, у свиньи специфические железы находятся в «пятачке», у лошади — в пальцевом мякише железы стрелки, у овцы — межпальцевые железы и т. д.
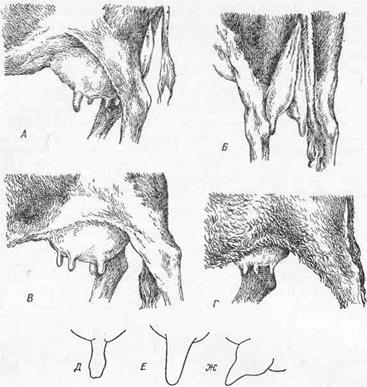 Рис. 8. Вымя и форма сосков крупного рогатого скота:
А — равномерно развитое («бедренное вымя»; Б—оно же, сзади; В — равномерное «брюшное вымя»; Г — примитивное вымя; Д — цилиндрический (мясной) сосок; Е— конический сосок; Ж — сосок, расширенный у основания.
Рис. 8. Вымя и форма сосков крупного рогатого скота:
А — равномерно развитое («бедренное вымя»; Б—оно же, сзади; В — равномерное «брюшное вымя»; Г — примитивное вымя; Д — цилиндрический (мясной) сосок; Е— конический сосок; Ж — сосок, расширенный у основания.
|
СТРОЕНИЕ ВЫМЕНИ
Вымя — uber —крупного рогатого скота (рис. 8) простое, расположено в лонной области между бедрами.
Снаружи вымя одето кожей, которая у животных, содержащихся на холоде, покрыта волосами. Каудальная поверхность вымени с ясно выступающими отвесными складками кожи и заметными линейными потоками волос называется молочным зеркалом. Под кожей вымени располагается поверхностная фасция (рис. 9—2), а под ней — глубокая фасция вымени(3), являющаяся продолжением желтой брюшной фасции. Глубокая фасция, отдавая на середине вымени два эластических листка, идущих от белой линии живота к основанию вымени, разделяет вымя на правую и левую половины и поддерживает его. Эти листки глубокой фасции составляют подвешивающую связку вымени (4). Поперечно, между сосками, вымя разделяется условно на переднюю и заднюю половины, то есть имеет четыре четверти, нерезко разграниченные между собой. Каждая четверть вымени имеет свои выводные протоки (7) и отдельный сосок. Иногда бывает шесть сосков. Чаще добавочные соски встречаются на задней половине вымени. Эти соски иногда функционируют.
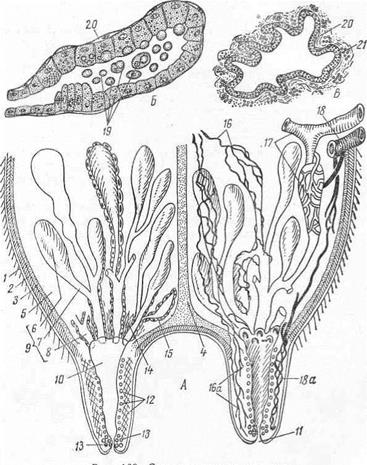 Рис. 9. Строение вымени коровы:
А—общая схема вымени в разрезе; Б — концевой отдел железы; В—крупный выводной проток; 1— кожа; 2 — поверхностная фасция; 3 — глубокая фасция; 4 — подвешивающая связка; 5 — строма; 6 — концевые отделы; 7 — мелкие выводные протоки; 8 - молочные ходы; 9 — паренхима; 10 — молочная цистерна; 11 — сосковый проток; 12 — гладкие мышечные клетки вокруг соска; 13— кольцевые мышцы, образующие сфинктер соскового канала; 14— пучки гладких мышц, сопровождающие крупные выводные каналы; /5 — миоэпителий, окружающий концевые отделы и выводные протоки; 16 — нервы; 16а — нервные окончания; /7 — артерия и ее ветвь, оплетающая концевой отдел железы; 18—вена вымени; 18а — венозное сплетение соска; 19 — элементы молока; 20 — миоэпителий; 21 — эпителий выводного протока.
Рис. 9. Строение вымени коровы:
А—общая схема вымени в разрезе; Б — концевой отдел железы; В—крупный выводной проток; 1— кожа; 2 — поверхностная фасция; 3 — глубокая фасция; 4 — подвешивающая связка; 5 — строма; 6 — концевые отделы; 7 — мелкие выводные протоки; 8 - молочные ходы; 9 — паренхима; 10 — молочная цистерна; 11 — сосковый проток; 12 — гладкие мышечные клетки вокруг соска; 13— кольцевые мышцы, образующие сфинктер соскового канала; 14— пучки гладких мышц, сопровождающие крупные выводные каналы; /5 — миоэпителий, окружающий концевые отделы и выводные протоки; 16 — нервы; 16а — нервные окончания; /7 — артерия и ее ветвь, оплетающая концевой отдел железы; 18—вена вымени; 18а — венозное сплетение соска; 19 — элементы молока; 20 — миоэпителий; 21 — эпителий выводного протока.
|
Железистая часть вымени — паренхима(9) построена по типу сложной альвеолярно-трубчатой железы и одета собственной соединительнотканной капсулой со скоплением жировых клеток и эластических волокон. От капсулы внутрь вымени направляется ряд пластинок и тяжей, разделяя его на отдельные железистые участки — дольки вымени. От междольковых соединительнотканных пластинок внутрь дольки идут нежные пучки, оплетающие конечные трубочки и альвеолы, или альвеолотрубки железы. Соединительнотканный остов вымени называется стромой или интерстицией. По ней проходят сосуды и нервы внутрь железы.
Стенка молочных альвеол состоит из: однослойного кубического эпителия, слоя миоэпителиальных (корзинчатых) клеток, базальной мембраны и периальвеолярной соединительной ткани, содержащей кровеносные и лимфатические капилляры и нервные волокна. Молочные железы секретируют по апокриновому типу, каждая секретирующая клетка выделяет сразу все составные части молока.
У секретирующих клеток мутная цитоплазма и округлое ядро и на свободной поверхности имеются микроворсинки.
Из альвеолотрубок (6) молоко переходит в тончайшие выводные протоки, выстланные однослойным кубическим эпителием, которые, соединяясь между собой, образуют видимые простым глазом молочные каналы (протоки), соединяющиеся в молочные ходы (в них эпителий становится двухслойным), которые, расширяясь около основания соска, открываются в полость — молочную цистерну (10).
Выводные протоки и концевые отделы молочной железы густо оплетаются сетью кровеносных капилляров (17, 18а) и нервными окончаниями (16а).
Сосок имеет молочную цистерну (10) и сосковый проток (11). Внутренний слой стенки молочной цистерны — слизистая оболочка — состоит из двухслойного призматического эпителия, слоя миоэпителия и собственной оболочки, снаружи от нее находятся пучки гладких мышечных волокон. Слизистая оболочка молочной цистерны образует много продольных складок, расправляющихся при наполнении цистерны молоком. Нижний конец молочной цистерны суживается и переходит в короткий сосковый проток (11), стенки его выстланы плоским многослойным эпителием. Гладкая мускулатура соска состоит из четырех слоев (12): продольного (глубокого), кольцевого, смешанного и радиального (поверхностного).
Кольцевой слой, сильно развиваясь вокруг соскового канала, образует сфинктер соска (13). Снаружи сосок одет кожей, в нем нет ни сальных, ни потовых желез, ни волос, но имеется большое количество нервных окончаний (16а).
Форма вымени и сосков у разных животных неодинакова. В известных случаях она связана с продуктивностью животных (рис. 10). У овцы и козы вымя состоит из правой и левой половин с одним соском на каждой половине (рис. 11—А). У крупного рогатого скота в каждой половине вымени имеется два соска с одним отверстием в каждом соске (Б). Вымя лошади имеет по одному соску на каждой половине, но в каждом соске две цистерны и два отверстия (В). Молочные железы (множественное вымя) свиньи состоят из 10—16, чаще 12 холмов со стольким же количеством сосков. В каждом соске по одной, чаще по две, молочной цистерне с сосковыми каналами.
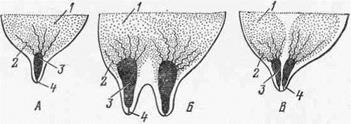 Рис. 11. Различие в строении молочных желез у мелких жвачных (А), коровы (Б), лошади (В).
1 — железистая масса; 2 — выводные протоки; 3 — молочная цистерна; 4 — сосковый проток.
Рис. 11. Различие в строении молочных желез у мелких жвачных (А), коровы (Б), лошади (В).
1 — железистая масса; 2 — выводные протоки; 3 — молочная цистерна; 4 — сосковый проток.
|
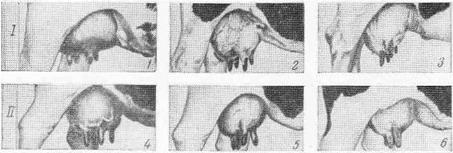 Рис. 10. Различные формы вымени рогатого скота (/):
/ — чашеобразное вымя; 2 — округлая форма вымени; 3 — козье вымя; II — формы округлого вымени; 4 — нормальное без пороков; 5— задние соски сближены; 6 — задние и передние соски сближены.
Рис. 10. Различные формы вымени рогатого скота (/):
/ — чашеобразное вымя; 2 — округлая форма вымени; 3 — козье вымя; II — формы округлого вымени; 4 — нормальное без пороков; 5— задние соски сближены; 6 — задние и передние соски сближены.
|
Изменение железистой ткани вымени в различные периоды жизни рогатого скота. Состояние железистой ткани вымени разных животных и даже у одного и того же животного в разные периоды его жизни, при разном уровне кормления бывает различным (рис. 12). До половой зрелости железистая ткань вымени не развивается. Развитие этой ткани происходит в тесной взаимосвязи с развитием соединительнотканного остова вымени. При этом увеличение паренхимы вымени вызывает относительное уменьшение его стромы, и, наоборот, уменьшение железистой ткани обусловливает некоторое увеличение соединительнотканного остова вымени. При наступлении половой зрелости животного кровеносные сосуды и нервы вымени сильно разрастаются и густо оплетают железистые элементы. Железистая и соединительная ткани начинают усиленно развиваться. Однако строма вымени развивается быстрее, чем паренхиматозные элементы, которые уже принимают форму альвеол. Только с наступлением беременности железистая
ткань начинает обгонять в развитии соединительнотканный остов; в некоторых отделах железы появляются в это время полости, формируется их стенка; железа по строению приближается к состоянию, обеспечивающему ее функцию.
К концу беременности молочные железы полностью развиваются и начинают продуцировать молозиво.
Наибольшего развития и наивысшей функциональной деятельности молочные железы достигают в период лактации. В это время просветы альвеолотрубок расширяются, заполняются секретом, а соединительнотканный остов их относительно уменьшается (А). К концу лактационного периода в железе наблюдаются обратные явления: альвеолотрубки постепенно прекращают свою работу, сжимаются, а соединительнотканный остов разрастается, и при хорошем питании животного в нем образуются жировые отложения (Б).
С наступлением новой беременности в вымени вновь начинает преобладать железистая ткань.
У высокомолочных коров соединительнотканный остов нежен, а трубочки и альвеолы тесно прилежат друг к другу, занимая большую часть дольки вымени. У малопродуктивных коров, наоборот, строма развита сильно, железистые же образования выражены слабее и лежат на значительном расстоянии одно от другого, отделяемые толстыми соединительнотканными прослойками.
У телок и старых коров строма также преобладает над паренхимой (В). У хорошо упитанных животных строма увеличивается за счет отложения жира (жировое вымя) (Г).
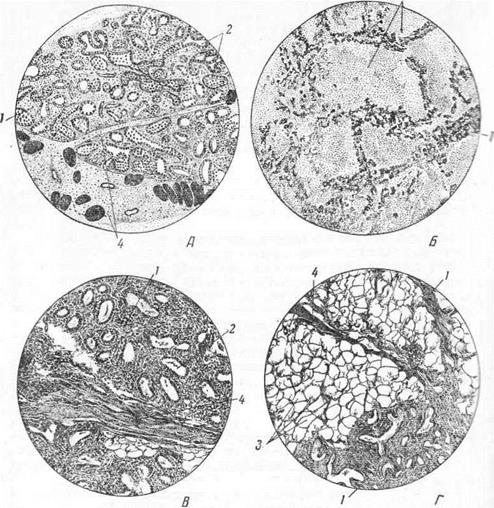 Рис. 12. Молочная железа коровы в состоянии лактации (А) и сухостоя (Б) и молочная железа телки при обычном (В) и обильном (Г) кормлении:
1 — внутридолькоаая соединительная ткань; 2 — трубчатые альвеолы; 3 — жировые клетки; 4 междольковая соединительная ткань.
Рис. 12. Молочная железа коровы в состоянии лактации (А) и сухостоя (Б) и молочная железа телки при обычном (В) и обильном (Г) кормлении:
1 — внутридолькоаая соединительная ткань; 2 — трубчатые альвеолы; 3 — жировые клетки; 4 междольковая соединительная ткань.
|
СТРОЕНИЕ МЯКИШЕЙ
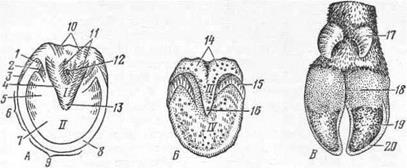
Мякиши— pulvini — бывают запястные (заплюсневые), пястные (плюсневые) и пальцевые (рис. 2). Наибольшее практическое значение имеют пальцевые мякиши лошади. Поэтому они подробнее и описываются.
Пальцевый мякиш лошади — pulvinus digitalis (рис. 13—I, III) — форма раздвоенного у основания треугольника, вклинивающегося своей вершиной в подошву копыта (II). Его задняя, более эластичная часть составляет подушку мякиша (10), а острая, более упругая, с массивным роговым слоем передняя часть — стрелку (11). Острый конец стрелки называют верхушкой (А—13). Выступающие па подошвенной поверхности роговые валики пальцевого мякиша это ножки стрелки (11), а находящееся между ними углубление — межножковый желоб (12). С внутренней стороны рогового слоя мякиша межножковой борозде соответствует гребень стрелки.
 |
Пальцевый мякиш, являясь производным кожного покрова, состоит из трех слоев: эпидермиса, основы кожи и подкожного слоя мякиша.
Рис. 13. Роговой слой копытной подошвы и пальцевого мякиша (А) и основа кожи копытной подошвы и пальцевого мякиша лошади (Б), роговой слой подошвы копытец и пальцевых мякишей крупного рогатого скота (В):
I— роговой слой пальцевого мякиша;II— роговой слой копытной подошвы;III— основа кожи пальцевого мякиша;IV— основа кожи копытной подошвы;1— угол копытней стенки;2—заворотная часть копытной стенки;3— задние части копытной стенки;4- боковые борозды;5 — ветви копытной подошвы;6— боковые части копытной стенки;7— тело копытной подошвы;8— подошвенный край копытной стенки;9— зацепная часть копытной стенки;10— подушка мякиша;11— ножки стрелки;12— межножковый желоб;13— верхушка стрелки;14— основа кожи подушки мякиша:15— основа кожи заворотной части копытной стенки;16— основа кожи стрелки пальцевого мякиша;17— висячие (2 и 5) пальцы;18— пальцевый мякиш;19 — копытцевая подошва;20— копытцевая стенка.
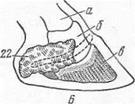 Рис. 14. Палец грудной конечности лошади:
Рис. 14. Палец грудной конечности лошади:
|
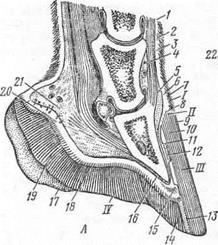 А — в сагиттальном разрезе; Б — положение мякишного хряща; I — копытная кайма; II — копытный венчик; III — копытная стенка; IV — копытная подошва; 1— эпидермис; 2 — основа кожи; 3—подкожный слой; 4 — сухожильное окончание общего пальцевого разгибателя; 5 — подкожныйслой копытной каймы и копытного венчика; 6 — основа кожи копытной каймы и копытного венчика; 7 — эпидермис копытной каймы и направление, в которомопускается роговой слой (глазурь) с каймына венчик и стенку; 8 — эпидермис копытноговенчика; 9 — глазурь копытной стенки;10 — трубчатый рог и направление, в котором он опускается с венчика на стенку копыта; 11 — листочковый рог копытной стенки; 12 — листочковый слой основы кожи копытной стенки; 13 — белая линия; 14 — роговой слой копытной подошвы; 15 — основа кожи копытной подошвы; 16 — надкостница; 17 — роговой слой стрелки пальцевого мякиша; 18 — основа кожи стрелки пальцевого мякиша; 19 — роговой слой подушки пальцевого мякиша; 20 — основа кожи подушки пальцевого мякиша;
А — в сагиттальном разрезе; Б — положение мякишного хряща; I — копытная кайма; II — копытный венчик; III — копытная стенка; IV — копытная подошва; 1— эпидермис; 2 — основа кожи; 3—подкожный слой; 4 — сухожильное окончание общего пальцевого разгибателя; 5 — подкожныйслой копытной каймы и копытного венчика; 6 — основа кожи копытной каймы и копытного венчика; 7 — эпидермис копытной каймы и направление, в которомопускается роговой слой (глазурь) с каймына венчик и стенку; 8 — эпидермис копытноговенчика; 9 — глазурь копытной стенки;10 — трубчатый рог и направление, в котором он опускается с венчика на стенку копыта; 11 — листочковый рог копытной стенки; 12 — листочковый слой основы кожи копытной стенки; 13 — белая линия; 14 — роговой слой копытной подошвы; 15 — основа кожи копытной подошвы; 16 — надкостница; 17 — роговой слой стрелки пальцевого мякиша; 18 — основа кожи стрелки пальцевого мякиша; 19 — роговой слой подушки пальцевого мякиша; 20 — основа кожи подушки пальцевого мякиша;
21 — подкожный слой подушки пальцевого мякиша; 22 — мякишный хрящ; а — путовая кость; б — венечная кость; в— копытная кость.
Эпидермис пальцевого мякиша — безволосый, имеет роговой, блестящий, зернистый и ростковый слои. В нем находятся трубчатые, свернутые клубком железы, выделяющие секрет жирового характера (10—13). В основе кожи пальцевого мякиша хорошо развиты сосочковый и сетчатый слои (14—16). Подкожный слой пальцевого мякиша от подкожного слоя кожного покрова отличается тем, что в нем в большом количестве присутствует жировая и эластическая ткани. Особенно много ее в подушке мякиша, выполняющей преимущественно рессорную функцию. Своеобразным видоизменением подкожного слоя пальцевого мякиша лошади являются парные (латеральный
и медиальный) мякишные хрящи, прочно сросшиеся с подушкой мякиша. Форма их в виде выпуклых пластинок и одним краем они плотно прирастают к копытной кости (рис. 14—Б—22). У старых животных, особенно у тяжеловозов, при неправильной их эксплуатации эти хрящи иногда окостеневают. Мякишные хрящи, охватывая с боков подушку мякиша, доходят проксимальным краем почти до половины венечной кости и хорошо прощупываются с волярной поверхности конечности. С близлежащими костями (копытной, челночной, венечной и путовой) хрящи соединены связками.
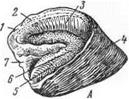
У рогатого скота и свиньи пальцевые мякиши стрелок не имеют, а подушки мякиша у них выражены хорошо (рис. 13—В—18). В остальном пальцевые мякиши рогатого скота и свиньи имеют то же строение, что и у лошади. У собаки пальцевые мякиши имеются на каждом пальце. Они сильно наплывают на коготь с подошвенной стороны.
 СТРОЕНИЕ КОПЫТА И РОГА
СТРОЕНИЕ КОПЫТА И РОГА
Копыто — ungula — лошади делят на копытную кайму, копытный венчик, копытную стенку и копытную подошву (рис. 14).
 Копытная кайма в виде узкой полосы, около 0,5 см ширины, составляет переход от волосатой кожи конечности к безволосой ее части (А—I). Копытный венчик шириной около 1,5 см располагается полукольцом ниже копытной каймы, составляя проксимальный край безволосой поверхности копыта (А— II). На его роговом слое с внутренней стороны имеется венечный желоб (рис. 15—2), которому на основе кожи копытного венчика соответствует венечный валик (Б—10). Копытная стенка (рис. 14—А—III) — наиболее массивная часть копыта. Составляет переднюю и боковую поверхности копыта, частично заходит на подошвенную поверхность копыта и делится на непарную зацепную (рис.
Копытная кайма в виде узкой полосы, около 0,5 см ширины, составляет переход от волосатой кожи конечности к безволосой ее части (А—I). Копытный венчик шириной около 1,5 см располагается полукольцом ниже копытной каймы, составляя проксимальный край безволосой поверхности копыта (А— II). На его роговом слое с внутренней стороны имеется венечный желоб (рис. 15—2), которому на основе кожи копытного венчика соответствует венечный валик (Б—10). Копытная стенка (рис. 14—А—III) — наиболее массивная часть копыта. Составляет переднюю и боковую поверхности копыта, частично заходит на подошвенную поверхность копыта и делится на непарную зацепную (рис.
13—А—9), парные боковые передние (6),
боковые задние (3), заворотные (2) части и заворотные углы (1). На копытной стенке, доходящей до дистального конца конечности, различают также подошвенный край (8). На копытной подошве (А—II), непосредственно опирающейся о почву, различают тело (7) и подошвенные ветви (5), между которыми вклинивается пальцевый мякиш, отделяющийся от подошвенных ветвей и заворотных частей копытной стенки боковыми бороздами.
Гистологическое строение копыта имеет много общего со строением кожи, но существуют и некоторые отличия, связанные с функциональным различием их.
 Копытная кайма (рис.14—А—I) состоит из эпидермиса, основы кожи и подкожного слоя. Эпидермис имеет роговой, зернистый и ростковый слои. Роговой слой безволосый. Опускаясь на копытную стенку, он формирует тонкий блестящий слой —глазурь копытной стенки (А—9; 16—1). Основа кожи состоит из сосочкового и сетчатого слоев (рис. 14—А—6). Сосочки сосочкового слоя опущены книзу. Подкожный слой каймы развит незначительно (А—5).
Копытная кайма (рис.14—А—I) состоит из эпидермиса, основы кожи и подкожного слоя. Эпидермис имеет роговой, зернистый и ростковый слои. Роговой слой безволосый. Опускаясь на копытную стенку, он формирует тонкий блестящий слой —глазурь копытной стенки (А—9; 16—1). Основа кожи состоит из сосочкового и сетчатого слоев (рис. 14—А—6). Сосочки сосочкового слоя опущены книзу. Подкожный слой каймы развит незначительно (А—5).
Копытный венчик (А—II) также имеет эпидермис, основу кожи и подкожный слой. Эпидермис безволосый и состоит из рогового, зернистого и росткового слоев. Роговой слой очень толстый, трубчатого строения, наиболее крепкий в копыте и почти непроницаем для воды. Растет он по направлению к подошве копыта, формируя трубчатый рог стенки копыта (рис. 16—2).
На внутренней поверхности рогового слоя эпидермиса есть углубление — венечный желоб (рис. 15 — А — 2). Основа кожи копытного венчика (рис. 14—А—6; 15—Б—10) имеет сосочковый и сетчатый слои. Сосочки в этой части копыта наиболее длинные, опущены вниз, вследствие чего и роговой слой копытного венчика растет вниз. В основе кожи венчика разветвляется огромное количество сосудов и нервов, обеспечивающих
прекрасное кровоснабжение всего этого участка и хорошее ощущение неровностей почвы, на которую ступает копыто. Подкожный слой копытного венчика (рис. 14—А—5) вместе с основой кожи венчика образует венечный валик (рис. 15—Б—10).
Копытная стенка (рис. 14—III) по строению значительно отличается от кожного покрова. Эпидермис ее лишен волос и состоит из росткового и рогового слоев. Зернистого и блестящего слоев в нем нет (С. Н. Кревер). Ростковый слой эпидермиса основной массой располагается в проксимальной ее части, на границе с венчиком. Продуцируемый нм рог в форме листочков — листочковый рог (рис. 16—4). В ростковом слое эпидермиса копытной стенки находятся не цилиндрические клетки, а кубические, и роговые листочки
не являются их производными. Предполагают, что кубические клетки способствуют сползанию сверху вниз рогового слоя, образующегося на границе венчика и копытной стенки. Цвет листочкового рога белый. Его конечный участок на копытной подошве вместе с внутренним пластом трубчатого рога формирует белую линию. По ее расположению определяют, где при ковке животного надо забивать гвозди, чтобы они шли в трубчатый рог (латерально от белой линии), а не в основу кожи копытной стенки, богатой сосудами и нервами. Таким образом, копытная стенка имеет три роговых слоя. Листочковый рог (4) находится под трубчатым рогом, идущим от копытного венчика, а трубчатый рог (2) прикрывается глазурью (1), спускающейся с копытной каймы. При этом глазурь у старых животных и у животных, содержащихся в плохих условиях, обычно разрушается, и тогда наружным слоем копытной стенки остается трубчатый рог.
 Основа кожи копытной стенки вместо сосочкового слоя имеет листочковый слой (6), каждый листочек которого представляет собой как бы слияние ряда сосочков. Между листочками основы кожи копытной стенки и продвигаются роговые листочки ее эпидермиса. На поверхности каждого листочка с обеих сторон располагается ряд
Основа кожи копытной стенки вместо сосочкового слоя имеет листочковый слой (6), каждый листочек которого представляет собой как бы слияние ряда сосочков. Между листочками основы кожи копытной стенки и продвигаются роговые листочки ее эпидермиса. На поверхности каждого листочка с обеих сторон располагается ряд
более мелких вторичных листочков. Это увеличивает поверхность всех листочков до одного квадратного метра, чем обеспечивается прочное соединение листочкового слоя основы кожи копытной стенки с листочковым рогом ее. Сетчатый слой основы кожи копытной стенки (7) из-за отсутствия подкожного слоя непосредственно соприкасается с копытной костью. Его внутренняя поверхность, срастаясь с костью, образует периостальный слой копытной
стенки. Такое прочное соединение основы кожи копытной стенки с копытной костью обеспечивает при действии мышц на копытную кость одновременное и сочетанное движение ее вместе с копытом.
Подкожного слоя копытная стенка не имеет. Основа кожи копытной стенки богата нервами и сосудами. При ковке животного гвозди не должны попадать в этот слой.
Копытная подошва (рис. 13—А—II) также не имеет подкожного слоя, что способствует прочному сцеплению основы кожи копытной подошвы с копытной костью. Эпидермис копытной подошвы (рис. 14—А—14), состоящий из рогового, зернистого и росткового слоев, без волос, но обладает мощным трубчатым роговым слоем, по прочности, однако, уступающим трубчатому рогу копытной стенки. Его поверхностные слои представляют крошковатую массу, которая постепенно отпадает, а в случае необходимости ее счищают. Основа кожи копытной подошвы (А—15) состоит из сосочкового и сетчатого слоев. Внутренняя поверхность сетчатого слоя образует периостальный слой, имеющий то же значение, что и соответствующий слой копытной стенки.
Роговые слои всех частей копыта и пальцевого мякиша составляют роговой башмак. Остальные части эпидермиса, а также основа кожи и подкожного слоя пальцевого мякиша и копыта в состав рогового башмака не входят. Различные отклонения в содержании животных отражаются на росте рогового башмака и вызывают его деформацию.
Копытца рогатого скотаи свиньи подобны копыту лошади, разделенному пополам (рис. 13—В), не имеют заворотных частей, а подошва их, особенно у мелкого рогатого скота, развита сильнее.
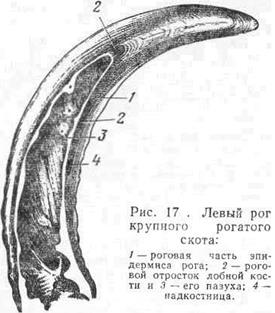
Строение рога. Рога располагаются на костных роговых отростках лобных костей и состоят из двух слоев — основы кожи и эпидермиса. У рогов различают корень, тело и верхушку (рис. 17). Основа кожи рога срастается с периостом роговых отростков лобных костей. Она имеет сосочковый и сетчатый слои. Сосочковый слой в различных участках его выражен неодинаково. У основания рога сосочки низки и расположены часто, дальше постепенно повышаются, располагаются реже и наклонно к верхушке рога, где находятся самые высокие сосочки. Эпидермис рога имеет те же слои, что и эпидермис копыта. Его ростковый слой продуцирует очень прочный трубчатый роговой слой. На росте рогового слоя отражаются различные условия существования животного — усиление или ослабление кормления, здоровое или болезненное состояние, нормальная или ослабленная функция росткового слоя. Усиленная функция росткового слоя ведет к появлению особых наплывов, колец на рогах. Ослабление в развитии рога делает его стенки более тонкими, образует перехват, или кольцо, на роге. У коров каждая стельность сопровождается появлением такого кольца. У многих животных кольца выражены по всей поверхности рога, у крупного рогатого скота, они ясно заметны лишь у корня рога. Верхушка рога у них бывает обычно гладкой и острой.
И.Т КРУГЛИКОВА
АНТИЧНАЯ АРХЕОЛОГИЯ

Допущено
Министерством высшего
и среднего специального
образования СССР
в качестве учебного пособия
для студентов
высших учебных заведений,
обучающихся по специальности
„История"
МОСКВА ВЫСШАЯ ШКОЛА 1984
ПРЕДИСЛОВИЕ
Основой настоящей книги послужил спецкурс, который читался в течение многих лет на историческом факультете МГУ для студентов, специализирующихся по археологии.
Широкий размах археологических исследований античных памятников на территории СССР и за рубежом способствовал накоплению массового бытового материала, что в сочетании со свидетельствами письменных и других видов источников позволяет дать исчерпывающую характеристику истории античного общества и €го материальной культуры, воссоздать достоверную картину жизни не только греков и римлян, но и многих других народов, на которых распространялось влияние греко-римской культуры.
Античные памятники, открытые на территории СССР, в районах, примыкающих к побережью Черного и Азовского морей, на Кавказе и в Средней Азии, в данном пособии впервые рассматриваются наряду с классическими античными памятниками Средиземноморья. Автор стремился выявить следы влияния античной культуры на народы, которые в ходе их исторического развития оказывались связанными прямо или опосредствованно с греками и римлянами либо с эллинистическими государствами Востока.
Античная культура, сложившаяся на территории рабовладельческих государств Греции и Рима в I тысячелетии до н. э. и в первые века нашей эры, сыграла ..значительную роль в истории человечества.К.Маркс называл античный период детством человеческого общества, которое прекраснее всего развилось в Греции1. Поэтому естественно, что основное внимание в книге уделяется анализу материальной культуры Греции и Рима. Античные памятники периферии этих государств рассматриваются под углом зрения степени распространения влияния этой культуры, и иллюстрации в книге характеризуют прежде всего наиболее важные стороны жизни греков и римлян на их территории. Прекрасные памятники античной культуры, найденные на территории нишей страны, представлены фотографиями художественных сосудов, статуэток, светильников, ювелирных украшений, стеклянных изделий и других произведений греческих и римских ремесленников. Все иллюстрации помещены в конце книги.
1 См.: Маркс К-, Энгельс Ф. Соч 2-е изд., т. 12, с. 737—738.
ВВЕДЕНИЕ
Античная археология — это археология греко-римских государств Средиземноморья и побережья Черного моря, в более широком понимании — районов, где находились поселения греков и римлян. Греческая культура в эпоху эллинизма распространилась далеко на Восток вплоть до Индии и Средней Азии, а в эпоху римской империи в ареал влияния античной культуры попадают обширные территории римских провинций от Испании до Передней Азии и от Британии до Африки.
Хронологически античная археология изучает период I тысячелетия до н. э. — 1-ю половину I тысячелетия н. э., когда в Средиземноморье сложились и развивались рабовладельческие государства, имевшие письменность, изобразительное искусство, развитую торговлю.
Античная археология тесно связана с данными письменных источников (сочинениями античных авторов, эпиграфикой), нумизматикой, архитектурными памятниками, памятниками искусства и т. д.
Термин «археология» происходит от греческого «архайос логос», что означает: наука о древностях. Он появился впервые в античном мире. Греческий философ Платон (427—347 гг. до н. э.) этим термином называл науку о древности (о старине). Позднее писатели I в. до н. э. и I в. н. э. — Диодор Сицилийский, Страбон археологией называли догомеровский, т. е. доисторический, период. Дионисий Галикарнасский назвал свою историю Римского государства «Римская археология», а Иосиф Флавий, написавший «Историю иудейского народа», называл ее иудейской археологией. У римлян термину «археология» соответствовал термин antiquitates, т. е. древности.
Поскольку письменные источники неодинаково полно освещают различные периоды истории и дают мало сведений о жизни простого народа — непосредственного производителя материальных благ, археологические источники восполняют эти пробелы. Они позволяют осветить историю материальной жизни народов. Памятники античного времени благодаря сочетанию с монетными и эпиграфическими находками дают возможность устанавливать довольно точные датировки археологических слоев и сооружений. При этом они датируют и широкий круг памятников, находимых на территории окружавшего античные государства варварского мира.
Археологам приходится изучать различные типы памятников античного времени: большие и малые города, сельские поселения,
отдельные сельские усадьбы или виллы, крепости, фортификационные сооружения, некрополи, включающие курганы, склепы, каменные ящики и простые грунтовые могилы. Методика раскопок этих объектов имеет свои специфические особенности. Редко встречаются античные города, оставленные жителями в результате единовременной катастрофы, как, например, Помпеи, засыпанные пеплом, или Геркуланум, залитый 15-метровым слоем лавы при извержении Везувия в 79 г. В пустынных районах Сирии, Ирана, Афганистана и других восточных государств, а также на территории советских Среднеазиатских республик встречаются руины городов, заброшенных жителями в связи с изменением режима рек или разрушением сети ирригационных сооружений. Иногда города преднамеренно разрушались завоевателями и больше не восстанавливались, как, например, Олинф на Халкидском полуострове, разрушенный Филиппом II в 349—348 гг. до н. э. Но чаще всего города существовали в течение многих столетий, благодаря чему их культурные слои сильно повреждены различными ямами, поверхность слоев многократно нивелировалась, что сопровождалось перемещением больших масс грунта. Это чрезвычайно затрудняет изучение стратиграфии античных городов, требует вскрытия больших площадей и тщательной фиксации различных грунтовых ям и наслоений.
В настоящее время при раскопках и разведках античных памятников широкое распространение получила аэрофотосъемка.
В 1925—1932 гг. французский ученый А. Пуадебар провел с помощью аэрофотосъемки большие археологические изыскания в Сирии и Ливане для изучения римского лимеса — системы пограничных укреплений и великого караванного пути, связывавшего Римскую империю со странами Востока. Ученый вел наблюдения над рельефом местности и над ее растительным покровом. Позднее он нсследовал гавани Тира и Сидона, сочетая аэрофотосъемку с подводными работами и раскопками на берегу.
С помощью аэрофотосъемок итальянский ученый Д. Адаместеану установил границы размежевания земельных владений Мета-понта. На территории Франции сплошная аэрофотосъемка целых районов дала возможность выявить систему расположения и даже планы античных усадеб.
Наряду с аэрофотосъемкой в античной археологии стали применять геофизические методы разведок. Особенно успешно их применял итальянский инженер М. Леричи для выявления этрусских склепов. Сейчас с помощью геофизических методов научились снимать планы нераскопанных каменных зданий, выявлять скрытые в земле кладки и пустоты погребальных памятников. Начала развиваться подводная археология. Все эти методы исследований античных памятников успешно применяются и в СССР. Кроме того, при обработке находок в лабораториях советские ученые используют методы палеонтологии, палеозоологии, палеоботаники, петрографии, применяют данные рентгенографии, химических и спектральных анализов. Это углубляет исследования и позволяет лучше понять жизнь древнейшего населения нашей страны.
Дата добавления: 2014-12-17; просмотров: 2575;