ТЕМА 6. ЦИТОПЛАЗМА КЛЕТКИ, ЕЁ КОМПОНЕНТЫ
Энергия, которая высвобождается в процессе расщепления молекул АТФ, используется для синтеза необходимых организму соединений, поддержания определенной температуры тела, обеспечения других процессов жизнедеятельности. Реализуется этот процесс путем взаимодействия молекул АТФ с ферментами, для которых они служат активаторами.
V. Биологически активные вещества – это органические соединения разной химической природы, способные влиять на обмен веществ и превращения энергии в живых существах. Одни из них регулируют процессы
метаболизма, роста и развития организмов, другие служат средством влияния на особей своего или других видов. К биологически активным веществам относят ферменты, витамины, гормоны, нейрогормоны, факторы роста, фитогормоны, антибиотики и т. п.
1. Витамины – низкомолекулярные органические соединения различной химической природы, необходимые для обеспечения процессов жизнедеятельности всех живых организмов. Витамины участвуют в обмене веществ и превращении энергии в основном в качестве компонентов сложных ферментов. Суточная потребность человека в витаминах составляет миллиграммы, а иногда и микрограммы. Известно приблизительно 20 разных витаминов и витаминоподобных соединений.
Основной источник витаминов для человека – продукты питания. Некоторые витамины в незначительном количестве синтезируются в организмах человека и животных из веществ-предшественников – провитаминов (так, витамин D образуется в коже человека под воздействием ультрафиолетового облучения) или симбиотическими микроорганизмами (в частности, в кишечнике человека бактерии синтезируют витамины К, В6, В12).
Когда еще не были известны химическое строение и механизм действия витаминов, их помечали буквами латинского алфавита – А, В, С, D др. Теперь применяют и химические названия (например, витамин С акже называют аскорбиновой кислотой). Витамины делят на две группы: водорастворимые (группа В, С и др.) и жирорастворимые (А, D, К, Е).
При недостатке определенных витаминов, их полном отсутствии, а также избытке развиваются опасные для здоровья человека и животных заболевания. Например, при недостатке определенных витаминов в организме возникает гиповитаминоз, при их полном отсутствии – авитаминоз, а при избытке – гипервитаминоз (например, избыточное содержание витамина А в организме человека, превышающее норму в 20–30 раз, вызывает отравление). Явления гипо- и авитаминоза могут проявляться и в результате нарушения обмена веществ. Это так называемая вторичная витаминная недостаточность, связанная с невосприятием организмом определенных витаминов.
2. Гормоны – органические вещества, регулирующие процессы обмена веществ и превращений энергии в организме животных и человека. Их производят железы внутренней и секреции (эндокринные железы). Именно секреция в специфических анатомических образованиях отличает гормоны от других регуляторных веществ, которые секретируются отдельными клетками, не оформленными в виде желез. К последним относят, в частности, нейрогормоны –вещества синтезирующиеся в некоторых нервных клетках, и потом поступающие в кровь(например, вазопрессин и окситоцин синтезируются в клетках мозга, оттуда поступают в эндокринную железу гипофиз, а уже из нее – в кровоток). Механизмы действия различных гормонов и нерогоромнов очень сильно различаются, но их объединяет общая функция: регуляция жизнедеятельности на уровне всего организма. Под контролем гормонов и нейрогормонов происходят все этапы индивидуального развития человека и животных и все процессы их жизнедеятельности. Если определенные гормоны производятся в недостаточном количестве или не производятся вообще, наблюдают нарушения развития и обмена веществ разной степени тяжести. Избыточная выработка определенных гормонов также негативно влияет на организм. Так, при недостаточном образовании в организме ребенка гормона роста развивается карликовость,а при избыточном – гигантизм.
По сравнению с работой нервной системы, которая обеспечивает передачу сигналов на значительные расстояния за короткое время, действие гормонов отличается меньшей скоростью, но производит более длительный эффект. Характерные особенности действия гормонов и нейрогормонов: дистанционность, высокие биологическая активность и специфичность действия, короткое время существования.
3. Нейромедиаторы – биологически активные вещества, которые обеспечивают межклеточные взаимодействия в нервной системе. В частности, они участвуют в передаче нервного импульса между двумя нейронами или между нейроном и мышечной клеткой. Их производят нервные и рецепторные клетки. Молекулы медиатора выделяются в пространство синаптической щели. Роль медиаторов могут исполнять различные вещества: ацетилхолин, норадреналин, и др. Гормон адреналин также выполняет функцию нейромедиатора.
4. Факторы роста клетки – полипептиды, стимулирующие или подавляют деление, дифференциацию, подвижность, метаболизм, другие проявления жизнедеятельности и гибель клеток. В отличие от гормонов, факторы роста продуцируют неспециализированные клетки, которые можно встретить во всех тканях.
5. Фитогормоны – регуляторные вещества растений, аналогичные по функциям гормонам животных. Однако по химической природе не имеют с последними ничего общего, так как возникли в ходе эволюции совершенно независимо. В отличие от животных, растения не имеют специальных органов, синтезирующих гормоны; вместе с тем отмечается большая насыщенность гормонами некоторых органов по сравнению с другими. Так, ауксинами богаче всего верхушечные меристемы стебля, гиббереллинами — листья, цитокининами — корни и созревающие семена. Фитогормоны обладают широким спектром действия. Фитогормоны регулируют многие процессы жизнедеятельности растений: прорастание семян, рост, дифференциацию тканей и органов, цветение, созревание плодов и т. п. Образуясь в одном органе (или его части) растения, фитогормоны обычно транспортируются в другой (или его часть).
6. Фитонциды— образуемые растениями биологически активные вещества, убивающие или подавляющие рост и развитие бактерий, микроскопических грибов, простейших. Характерными представителями фитонцидов являются эфирные масла, извлекаемые из растительного сырья промышленными методами.
7. Феромооны – продукты внешней секреции, выделяемых некоторыми видами животных и растений обеспечивающих химическую коммуникацию между особями одного вида. Если гормоны обеспечивают регуляцию на организменном уровне организации, то феромоны – на популяционно-видовом.
Приведенный выше список разновидностей биологически активных веществ далеко не полон, кроме того взгляды различных авторов, на их классификацию очень сильно разнятся. Само определение биологически активных веществ не позволяет установить сколько-нибудь четкие рамки для круга относящихся к ним соединений.
Литература:
Балан П.Г. Биология: Учебн. для общеобразоват. учебн. заведений: уровень стандарта, академический уровень / П.Г.Балан, Ю.Г.Верес,В.П.АПолищук; пер. с укр. – к,:Генеза, 2010. С. 58 – 70.
ТЕМА 5. ОБЩИЙ ПЛАН СТРОЕНИЯ КЛЕТОК. ПОВЕРХНОСТНЫЙ АППАРАТ. ЯДРО
План
I. История изучения клетки.
II. Методы клеточных исследований.
III. Общий план строения и разнообразие клеток живых организмов.
IV. Химический состав, структура и функции клеточных мембран.
V. Функции и особенности строения поверхностного аппарата клеток организмов различных царств живой природы.
VI. Строении и функция ядра эукариот. Кариотип.
VII. Наследственный материал прокариот.
Клетка – основная структурно-функциональная единица всех организмов, элементарная биологическая система. Изучению клеток посвящен раздел биологии – цитология.
I. Важнейшие вехи истории становления клеточной теории:
1665 г. Роберт Гук с помощью собственноручно сконструированного микроскопа рассмотрел оболочки клеток пробки и предложил термин «клетка».
1702 г. Антони ван Левенгук также с помощью микроскопов собственной конструкции открыл бактерий и одноклеточных животных (инфузорий и др.), описал эритроциты и сперматозоиды.
1828 г. Роберт Броун открыл ядро в клетках растений, а в 1833 г. предложил само название «ядро».
1830 г. Ян Евангелист Пуркине описал ядро в яйцеклетках курицы.
1838 г. Теодор Шванн, опираясь на труды Матиаса Шлейдена, сформулировал положения клеточной теории, основные из которых: все организмы состоят из клеток; клетки животных и растений сходны по строению и химическому составу.
1858 г. Рудольф Вирхов доказал, что клетки образуются не из бесструктурного межклеточного вещества, как считалось раньше, а в результате деления («всякая клетка – из клетки»).
II. Основные методы клеточных исследований:
1. Микроскопия. Основная проблема исследования структуры клеток связана с их мелкими размерами. Человеческий глаз обладает разрешающей способностью около 100мкм(1 мкм = 0,001 мм). Это означает, что две точки, расположенные на расстоянии менее чем 100 мкм друг от друга, кажутся одной расплывчатой точкой. Многие клетки и практически все внутриклеточные структуры имеют намного меньшие размеры. Для преодоления этой проблемы разработан ряд методов микроскопии:
- Световая микроскопия.В основе метода лежит способность оптических линз преломлять световые лучи и тем самым увеличивать образ объекта. Разрешающая способность микроскопов достигает 0,13—0,20 мкм, т. е. примерно в тысячу раз превышает разрешающую способность человеческого глаза. С помощью световых микроскопов, удается выявить многие детали внутреннего строения клетки — отдельные органеллы, клеточную оболочку. Основное ограничение световой микроскопии – длина волны световых лучей. Даже используя ультрафиолетовые лучи с длиной волны 250 – 400 нм невозможно добиться достаточного разрешения для рассмотрения деталей строения большинства внутриклеточных структур, так как их размеры еще меньше.
- Электронная микроскопия использует вместо лучей света поток электронов, а вместо линз – электромагниты. Разрешающая способность современных электронных микроскопов достигает 0,1 нм, поэтому с их помощью выявляют очень мелкие детали.
2. Цитохимические методы основаны на избирательном воздействии реактивов на определенные химические вещества цитоплазмы. Наиболее распространенным из них является дифференциальное окрашивание – на клеточный препарат воздействуют специальными красителями. окрашивающими строго определенные клеточные структуры (в силу их специфических химических свойств), а уже затем рассматривают под микроскопом. Это позволяет в деталях рассмотреть объекты, которые в обычных условиях практически полностью прозрачны и неразличимы.
3. Дифференциальное центрифугированиепозволяет разделить с помощью центрифуги содержимое клетки на отдельные разные по плотности составляющие и затем детально изучить химический состав каждой из них.
4. Метод меченых атомов, или авторадиография, помогает определить место и ход определенных физико-химических процессов в клетке. Для этого в нее вводят вещество, в котором один из атомов определенного элемента (углерода, фосфора и др.) замещен его радиоактивным изотопом. С помощью особых приборов, способных обнаруживать изотопы, можно проследить за перемещением (миграцией) в клетке этих веществ, их преобразованиями, выявить место и характер тех или иных биохимических процессов.
5. Метод клеточных культур – живые клетки содержат и размножают на искусственных питательных средах. Изменяя компоненты питательной среды, можно наблюдать, как те или иные соединения будут влиять на рост, размножение и другие свойства клеток.
III.По своей форме клеткиочень разнообразны. Организм человека, как и большинства животных, состоит из нескольких сотен разновидностей клеток. Значительное многообразие клеток присуще также и растениям.
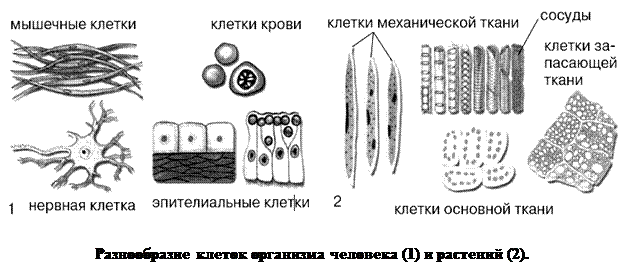 Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым закономерностям.
Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым закономерностям.
Существуют различные, иногда отчасти противоречивые, классификации структурных компонентов клеток. Приведем одну из наиболее традиционных и распространенных:

Цитоплазма –это все внутренне содержимоеклетки, включая ограничивающую ее извне мембрану – плазмолемму. Под плазмолеммой находится гиалоплазма (синонимы: матрикс цитоплазмы, цитозоль). Она представляет собой жидкую среду, содержащую до 90%, пронизанную молекулами фибриллярных белков. Смена конформации этих белков приводит к переходу гиалоплазмы из жидкого состояния в гелеобразное и наоборот. В гиалоплазму погружены различные органоиды и включения.
Органоиды, или органеллы – постоянные специализированные структуры в клетках живых организмов. Иногда клеточное ядро причисляют к двумембранным органоидам, а иногда рассматривают отдельно от цитоплазмы.Включения цитоплазмы — это необязательные компоненты клетки, появляющиеся и исчезающие в зависимости от интенсивности и характера обмена веществ в клетке и от условий существования организма. Это могут быть различные кристаллы, зерна, глыбки, капли и т. п. Включения могут просто плавать в цитоплазме, быть окруженными мембраной или располагаться внутри органоидов (например, в клеточном соке вакуоли растений).
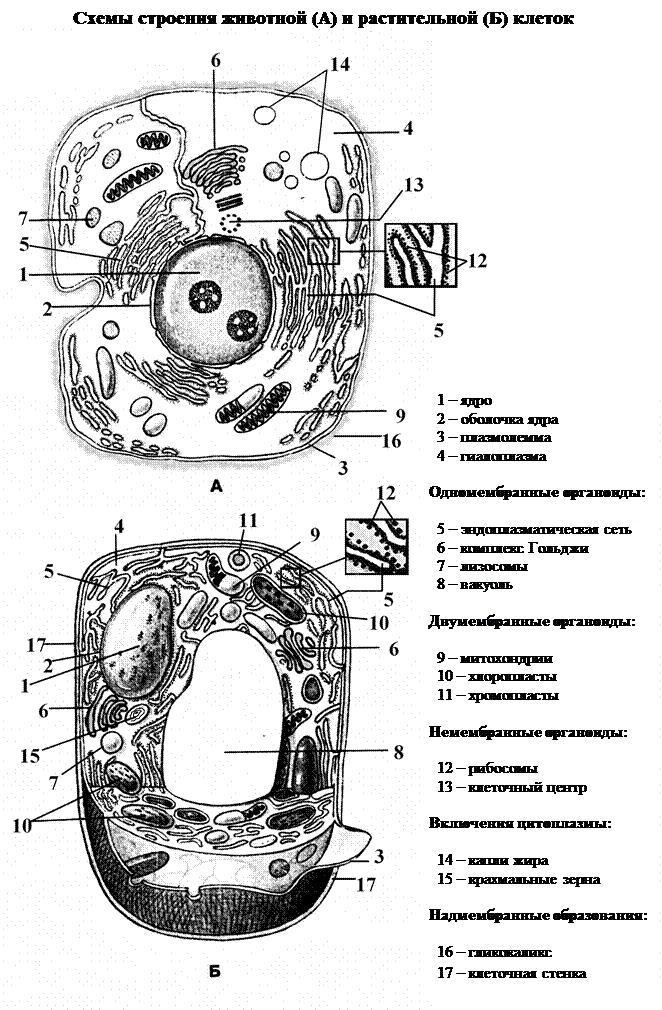
В зависимости от наличия ядра все организмы делят на два надцарства: Прокариоты и Эукариоты. Клетки прокариот, кроме того, что не имеют ядра, еще и довольно просто организованы. Клетки эукариот: грибов, растений и животных – организованы сложнее и обязательно имеют ядро (только в некоторых специализированных клетках ядро может на определенной стадии развития исчезать, например, в эритроцитах человека).
Снаружи от клеточной мембраны могут находиться надмембранные образования, различные по своей природе и функциям и специфичные для разных царств и подцарств живых организмов (у растений – целлюлозная клеточная стенка, выполняющая опорную и защитную функции, у многоклеточных животных – гликокаликс из различных углеводов и их соединений с белками и липидами, обеспечивающий взаимодействие клеток друг с другом, рецепцию и др.).
IV. Важнейшим структурным элементом клетки являются биологические мембраны. Мембрана, ограничивающая внутреннюю среду клетки, получила название плазмолемма, или плазматическая мембрана. Другие мембраны входят в состав различных органоидов. Толщина мембран, в зависимости от их типа, варьирует от 2–3 до 10 нм. Все разнообразные клеточные мембраны в общем имеют сходные химический состав и особенности организации.
Основу мембраны составляет липидный бислой. В нем молекулы липидов расположены в два слоя: их гидрофильные «головки» обращены к внешней и внутренней сторонам мембраны, гидрофобные «хвосты» – в глубь нее. Главный компонент бислоя – фосфолипиды, также в нем присутствуют гликолипиды и стероиды (у эукариот). Другая важная составляющая клеточных мембран – белки. Молекулы одних белков прилегают к липидному бислою изнутри или снаружи, их называют поверхностными, или переферическими, другие глубоко погружены в него (некоторые даже пронизывают насквозь) – интегральные, или внутренние белки. Углеводы мембраны связаны с ее белками и липидами, то есть являются частями молекул липо- и гликопротеидов. Они выступают над поверхностью мембраны. Особенно много их на наружной стороне плазмолеммы клеток животных, где они чувствуют в образовании ранее упоминавшейся надмембранной структуры – гликокаликса.
Такая модель строения биологических мембран получила название жидкостно-мозаичной. Название объясняется тем, что лишь приблизительно 30 % молекул липидов мембран крепко связаны с белками в единые комплексные соединения, а остальные – пребывают в жидком состоянии и способны свободно перемещаться. Благодаря этому мембраны быстро восстанавливаются после незначительных повреждений. Они могут легко сливаться между собой, растягиваться и сжиматься.

Плазмолемма – наружная мембрана клетки. Ее функции:
1. Барьерная – разделяет внешнюю среду и внутреннюю среду клетки, не давая им свободно смешиваться. В противном случае жизнь была бы невозможна. Также плазмолемма обеспечивает защиту от проникновения чужеродных объектов. В частности, вирусы могут проникнуть только в те клетки, на мембране которых есть специфические рецепторы, взаимодействующие с соответствующими рецепторами вируса (по принципу «ключ-замок». Поэтому подавляющее большинство вирусов животных не могут заразить человека – они просто неспособны проникнуть через плазмолемму.
2. Транспортная. Для жизнедеятельности клетки необходим обмен веществ с внешней средой. Плазмолемма характеризуется полупроницаемостью: одни соединения могут быстро проходить через нее, другие – медленнее или не проходят вообще. Механизмы транспорта веществ через мембрану различны:
- Пассивный транспортидет по градиенту концентрации, то есть из зоны с высокой концентрацией вещества в зону с низкой его концентрацией. Он происходит без затрат энергии. В основе механизма – процесс диффузии. При простой диффузии молекулы транспортируемого вещества свободно проникают через липидный бислой Путём простой диффузии в клетку проникают газы (O2, N2) гидрофобные вещества и полярные маленькие молекулы (CO2, H2O, мочевина). Не проникают полярные относительно крупные молекулы (аминокислоты, моносахариды), заряженные частицы (ионы) и макромолекулы (ДНК, белки). Благодаря такой избирательной проницаемости возможно явление осмоса – диффузии растворителя (воды) в область большей концентрации растворенного вещества (сахаров, солей, которые не могут свободно покинуть клетку). Благодаря этому внутри клетки создается повышенное – осмотическое – давление. Оно достигает значительных величин в клетках растений обеспечивая тургор – их упруго состояние. Облегченная диффузия – проникновение через мембрану определенных молекул с помощью мембранных белков-переносчиков, пронизывающих мембрану. Эти белки пропускают через себя только специфические для каждого виды молекул. Таким образом через мембрану проходит большинство веществ. Облегченная диффузия происходит медленнее простой, так как число молекул белков-переносчиков ограничено.
- Активный транспортосуществляется против градиента концентрации. Он связан со значительными затратами энергии, аккумулированной в молекулах АТФ. Например, концентрация ионов калия внутри клетки выше, чем снаружи, а натрия наоборот. Благодаря этой разности концентраций в результате диффузии ионы натрия поступают в клетку, а калия – выводятся из нее. Но концентрация этих ионов в живой клетке и снаружи нее никогда не выравнивается, поскольку существует особый механизм, благодаря которому ионы натрия выходят («откачиваются») из клетки, а калия – поступают («закачиваются») в нее. Этот механизм, действующий благодаря специальному белку, получающему энергию от разложения АТФ, получил название калий-натриевого насоса.
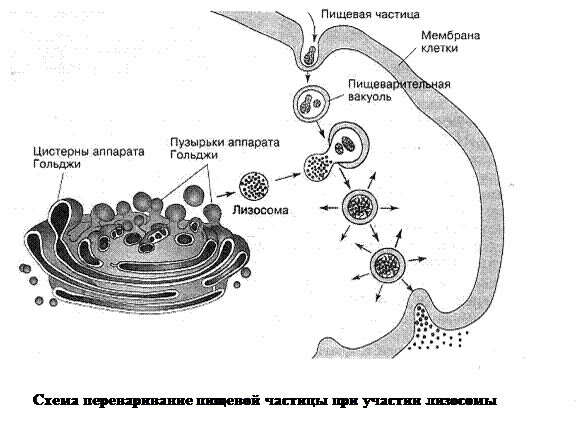
- Эндоцитоз –особый способ поглощения клеткой различных объектов, происходящий в ряд этапов: плазмолемма впячиватся захватывая внутрь цитоплазмы поглощаемый объект, затем образовавшийся канал смыкается позади объекта, в результате чего от плазмолеммы обособляется внутрь клетки мембранный пузырек, содержащий поглощенный объект внутри себя – пищеварительная вакуоль (не путать с вакуолью с клеточным соком растений). В случае, когда этот объект представляет собой твердую частицу, говорят о фагоцитозе, а если жидкость – о пиноцитозе. Эндоцитоз характерен характерен, в основном, для клеток животных. В частности, таким способом питаются простейшие. На фагоцитозе основан один из механизмов клеточного иммунитета человека.
- Экзоцитоз –процесс, повторяющий эндоцитоз в обратном направлении, служит для выделения из клетки продуктов жизнедеятельности: отходов, секретов желез и т. п.
3. Рецепторная функцияобусловлена локализацией на поверхности плазмолеммы клеточных рецепторов, воспринимающих различного рода раздражения: механические, химические, электрические и др. для обеспечения адекватной реакции отдельной клетки или многоклеточного организма в целом.
4. Передача возбуждения –плазмолемма является основным элементом в передаче сигналов по клеткам возбудимых тканей животных (нервной, мышечной и железистой).
5. Межклеточные взаимодействия.Плазмолемма клеток растений образуетканалы, проходящие сквозь поры целлюлозной клеточной стенки и сливающиеся с плазмолеммой соседних клеток. Внутри этих каналов находится гиалоплазма и трубки эндоплазматической сети, идущие из одной клетки в другую. Эти образования в виде межклеточных цитоплазматических тяжей называются плазмодесмы. Благодаря ним клетки всего растения формируют единое образование – симпласт. В отличие от растений, у клеток животных нет прямых цитоплазматических контактов с соседними, однако они соединяются десмосомами – участками, где плазмолеммы двух клеток прочно сцеплены друг с другом особыми белками. Это обеспечивает, в частности, прочное соединение клеток кожи, сердечной мышцы.
6. Образование надмембранных структур. Например, клеточная стенка растений, является продуктом ферментативного синтеза белков плазмолеммы, производящих волокна целлюлозы.
V. К поверхностному аппарату клеток относят плазмолемму, надмембранный комплекс и подмембранный комплекс. Строение плазмолеммы было рассмотрено выше.
Надмембранный комплекс, или надмембранные образованияпокрывают плазмолемму извне, образуя дополнительный барьер между клеткой и внешней средой. Природа и структура его различна у организмов разных царств живой природы:
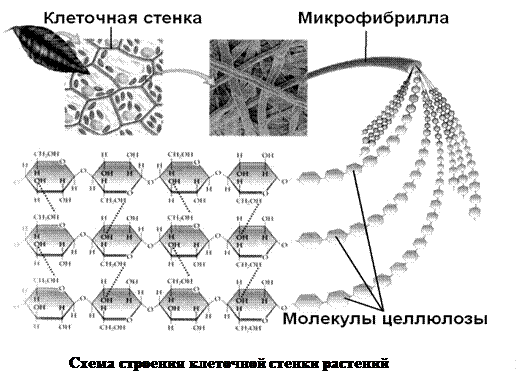
1. Клеточная стенка,иликлеточная оболочка растенийявляется продуктом выделения цитоплазмы. Она имеет следующий состав:
- целлюлоза – основной структурный компонент, откладывается в виде длинных пучков молекул – микрофибрилл, в совокупности образующих плотную решетку, окружающую клетку;
- гемицеллюлоза – аморфный компонент, представляющий собой смесь различных полисахаридов;
- пектины – полисахариды из остатков галактуровновой кислоты, в виде коллоида (взвести в воде), заполняют промежутки между микрофибриллами целлюлозы и молекулами гемицеллюлозы, увеличивая эластичность клеточной стенки;
- белки – ферменты, катализирующие расщепление целлюлозы, необходимое для роста клетки, а также процессы модификации клеточной стенки за счет откладывания в ней либо на ее поверхности лигнина, суберина и других веществ, происходящие в некоторых видах таканей, придавая им уникальные свойства (жесткость и прочность – волокном древесины, водонепроницаемость – клеткам пробки и др.).
- вода – реакционная среда для протекания вышеупомянутых ферментативных реакций, также изменение содержания регулирует прочность и эластичность клеточной стенки.
Аморфные вещества клеточной стенки поступают из глубины цитоплазмы в пузырьках комплекса Гольджи. Целлюлозные микрофибриллы синтезируются уже на поверхности плазмолеммы закрепленными в ней ферментами.
Немодифицированная (обычная) клеточная стенка полностью проницаема для воды и низкомолекулярных соединений. Она имеет поры, через которые проходят плазмодесмы, соединяющие все клетки растения.
Функции клеточной стенки:
- определяет форму клетки;
- опорная (особенно выражена у модифицированных оболочек механических тканей, например, волокон древесины);
- защитная – защищает клетку от разнообразных механических воздействий, вредителей и др.
- ограничивает увеличение объема клетки под действием осматического давления, которое в противном случае привело бы к разрыву плазмолеммы и гибели клетки, а благодаря клеточной стенки лишь создает тургор.
- транспортная – по клеточным стенкам транспортируется вода и растворенные в ней вещества, как к отдельным клеткам. так и в масштабе всего организма; стенки всех клеток растения соединены в общую систему – апопласт. В отличие от транспорта по симпласту через плазмодесмы, транспорт по апопласту возможен даже через мертвые ткани.
2. Клеточная стенка грибовсхожа с клеточной стен кой растений, но состоит не из целлюлозы, а из азотсодержащего полисахарида хитина.Несет аналогичные функции.
5. Клеточная стенка прокариот по своему строению разнообразна. Она может твердой либо иметь вид слизистой капсулы. Основной компонент клеточной стенки большинства бактерий – пептидогликан, или муреин(углеводно-пептидное соединение). У некоторых в ее состав входит особая дополнительная наружная мембрана, напоминающая плазмодлемму, и также обладающая полупроницаемостью, задерживая, в частности, молекулы многих антибиотиков. У цианобактерий в наружном слое клеточной оболочки находятся сократительные белки, обеспечивающие движения клеток путем скольжения и вращения.
6. Гликокаликс клеток животных, состоящий из углеводных частей молекул гликолипидов и гликопротеидов плазмолеммы уже рассматривался выше. Он обеспечивает непосредственную связь клеток с окружающей средой. В его состав входят рецепторные молекулы, способные воспринимать раздражители окружающей среды. Он также участвует в избирательном транспорте веществ (пропускает или не пропускает) молекулы, в зависимости от их размеров, заряда и т. п. В гликокалексе некоторых клеток локализированы ферменты обеспечивающие примембранное пищеварение – расщеплении соединений, которые расположены снаружи вблизи поверхности клетки. В итоге клетка потребляет эти продукты расщепления. Кроме того, гликокаликс обеспечивает межклеточные связи у многоклеточных животных и человека.
Подмембранный комплекс состоит из белковых микронитей и микротрубочек – элементов цитоскелета, локализованных под плазмолеммой и придающих ей дополнительную механическую прочность. Обычно эти структуры относят к немембранным органоидам, поэтому они будут более подробно рассмотрены в соответствующем разделе. Важными функциями подмембранного комплекса клеток являются обеспечение эндо- и экзоцитоза, движения (образование псевдоподий) и т. п. При этом микротрубочки и микронити удлиняются или укорачиваются.
В клетках многих одноклеточных животных (инфузорий, эвглен и т. п.) к подмембранным комплексам относится пелликула. Она состоит из структур, расположенных в уплотненном внешнем слое цитоплазмы. Так, у инфузорий в состав пелликулы входят уплощенные цистерны из органического вещества, которые вместе образуют мозаичную структуру. Внутри таких цистерн могут находиться дополнительные опорные комплексы из белков или пропитанных карбонатом кальция полисахаридных пластинок. Пелликула придает прочность оболочке клетки, обеспечивая относительное постоянство ее формы.
VI. Ядро – обязательная составляющая всякой эукариотической клетки, в нем сохраняется наследственная информация. Ядро регулирует процессы жизнедеятельности клеток. Лишь некоторые типы клеток эукариот лишены ядра. Это, в частности, эритроциты человека, ситовидные трубки высших растений. В таких клетках ядро формируется на начальных этапах развития, а потом разрушается. Потеря ядра сопровождается утратой способности клетки к делению.
Ядро выполняет две важнейшие функции:
- Является местом хранения и воспроизводства генетической информации, которая передается от материнской клетки дочерним в процессе клеточного деления;
- Ядро управляет всеми процессами жизнедеятельности клетки через синтез белков. В ядре генетическая информация перепереносится из ДНК в РНК, с помощью которых которой уже за пределами ядра синтезируются белки.
В клетках обычно есть лишь одно ядро, но некоторые клетки содержат от двух до нескольких тысяч ядер (инфузории, фораминиферы, некоторые водоросли, грибы, и др.). Зачем некоторым клеткам необходимо не одно ядро, а несколько или много? Дело в том, что каждому типу клеток присуще определенное постоянное соотношение между объемами ядра и цитоплазмы (ядерно-цитоплазматическое соотношение). Ведь ядро определенного объема может обеспечивать процессы биосинтеза белков лишь в соответствующем объеме цитоплазмы. Поэтому в клетках больших размеров или с повышенной интенсивностью обмена веществ часто находится от двух до нескольких тысяч ядер.
Форма ядра достаточно разнообразна. Чаще всего она сферическая или эллипсовидная, реже – неправильная (например, у некоторых типов лейкоцитов ядра имеют отростки). Размеры ядер варьируют от 1 мкм (некоторые одноклеточные животные, водоросли) до 1 мм (яйцеклетки некоторых рыб и земноводных). Ядро состоит из поверхностного аппарата и внутренней среды (матрикса). Поверхностный аппарат ядра образован двумя мембранами – внешней и внутренней, между которыми находится заполненное жидкостью щелевидное пространство шириной от 20 до 60 нм. В некоторых местах внешняя мембрана соединена с внутренней вокруг микроскопических отверстий – ядерных пор диаметром около 100 нм. Совокупность пор и таких белков называют поровыми комплексами. Поверхностный аппарат обеспечивает регуляцию транспорта веществ внутрь ядра и из него наружу через поровые комплексы. Он, например, распознает и сортирует эти соединения. Из цитоплазмы в ядро поступают белки, а из ядра в цитоплазму – разные типы молекул РНК и предшественники рибосом.
Ядерный матрикс – внутренняя среда ядра – состоит из ядерного сока, ядрышек и нитей хроматина. Ядерный сок (кариоплазма, или нуклеоплазма) по строению и свойствам напоминает гиалоплазму. В кариоплазме есть белковые фибриллы (нити) 2–3 нм толщиной. Они образуют особый внутренний скелет ядра, который соединяет различные структуры: ядрышки, нити хроматина, ядерные поры и т. п. Белки матрикса обеспечивают определенное пространственное расположение хроматина, а также влияют на его активность.
Хроматин – нитевидные структуры ядра, образованные в основном из белков и нуклеиновых кислот (ДНК и РНК). Участки хроматина неоднородны. Важнейший функциональный элемент хроматина – дезоксирибонуклеопротеид (ДНП) – комплекс ДНК с так называемыми гистоновыми белками, которые обеспечивают ее пространственное расположение, необходимое в тот или иной момент жизнедеятельности клетки. Так для синтеза РНК на матреце ДНК последняя должна быть полностью развернута. Если синтез не идет, гистоновые белки сворачивают ДНК в компактную суперспираль для экономии места. В одно и то же время одни участки ДНК развернуты, другие – свернуты. И только во время деления все молекулы ДНК ядра вместе с гистоновыми белками плотно свернута в компактные тела – хромосомы.
Ядрышки – плотные структуры, состоящие из комплексов РНК с белками, хроматина и гранул – предшественников рибосом. В ядре может быть от одного до многих ядрышек, которые формируются на особых участках хромосом. Во время деления клетки ядрышки исчезают вместе с ядерной оболочкой, а в период между двумя делениями – формируются снова. Функции ядрышек заключаются в образовании рРНК и предшественников рибосом, которые после формирования поступают в цитоплазму.
 Хромосомы, как уже было сказано выше, оформляются в виде различимых плотных тел только во время деления клетки (либо во время деления ядер без деления клетки). Длина их, в среднем, в среднем составляет 0,5–1,0 мкм, в то время как длина развернутых молекул ДНК – несколько сантиметров. Каждая хромосома состоит из двух продольных частей – хроматид, соединенных между собой в месте, названном зоной первичной перетяжки. Она разделяет хромосомы на два участка – плечи.
Хромосомы, как уже было сказано выше, оформляются в виде различимых плотных тел только во время деления клетки (либо во время деления ядер без деления клетки). Длина их, в среднем, в среднем составляет 0,5–1,0 мкм, в то время как длина развернутых молекул ДНК – несколько сантиметров. Каждая хромосома состоит из двух продольных частей – хроматид, соединенных между собой в месте, названном зоной первичной перетяжки. Она разделяет хромосомы на два участка – плечи.

В зоне первичной перетяжки имеется участок особого строения, который соединяет сестринские хроматиды – центромера. Здесь формируются белковые структуры – кинетохоры. Во время деления клетки к кинетохору присоединяются нити веретена деления, что обеспечивает упорядоченное распределение хромосом или отдельных хроматид между дочерними клетками (этот процесс будет подробнее рассмотрен в разделе, посвященном делению клеток). Некоторые хромосомы имеют еще и вторичную перетяжку, где расположены гены, отвечающие за образование ядрышек.
Каждая из хроматид содержит по молекуле ДНК со сходными наборами наследственной информации. Во время деления клетки хроматиды расходятся к дочерним клеткам, а в период между двумя делениями число хроматид снова удваивается. Это происходит благодаря способности молекул ДНК к самоудвоению (репликации).
Клетки каждого вида животных, растений, грибов имеют определенный набор хромосом. Совокупность признаков хромосомного набора (количество хромосом, их форма и размеры) называют кариотипом. Каждому виду присущ особый уникальный кариотип. Например, в неполовых клетках мухи-дрозофилы всего 8 хромосом (4 пары), человека – 46 (23 пары), у морских одноклеточных животных радиолярий – до 1600. Постоянность кариотипа обеспечивает существ существование видов. Специфический кариотип особей одного вида дает им возможность спариваться между собой и производить жизнеспособных потомков. При спаривании особей разных видов потомство не появляется вообще или же оно нежизнеспособно или бесплодно.
Хромосомный набор ядра может быть:
- гаплоидный (его условно обозначают 1n) – все хромосомы по строению отличаются одна от другой.
- диплоидный (2n) – каждая хромосома имеет парную, подобную по размерам и особенностям строения; их называют гомологичными; соответственно хромосомы, которые относятся к разным парам, не гомологичны друг другу.
- полиплоидный – количество гомологичных хромосом превышает две: триплоидный (3n), тетраплоидный (4n) и т. д.
У человека диплоидный набор хромосом.
У раздельнополых животных и двудомных растений у особей одного пола хромосомы одной из пар различаются между собой, тогда как у особей другого – они подобны. Это половые хромосомы, которые еще называют гетерохромосомами. Так, в хромосомном наборе женщины две Х-хромосомы, а мужчины – одна Х-хромосома и одна Y-хромосома. Хромосомы других пар, подобные у особей разных полов, называют неполовыми, или аутосомами. Естественно, если гомологичные аутосомы имеют подобный набор генов, то в Х- и Y-хромосомах он разный. Именно это различие и определяет развитие половых признаков.
У мух и млекопитающих разные половые хромосомы имеют особи мужского пола, а вот у бабочек, пресмыкающихся и птиц, наоборот, – женского. У некоторых животных особи разных полов имеют разное количество половых хромосом. Так, самки кузнечиков имеют 2 половые хромосомы, тогда как самцы – лишь одну.
Кариотип может изменяться вследствие хромосомных мутаций. Мутанты часто неспособны скрещиваться с теми, которые имеют нормальный кариотип, и оставлять плодовитых потомков. Кроме того, хромосомные мутации могут вызывать различные заболевания. В качестве примера можно привести болезнь Дауна. Мы уже упоминали, что человек имеет 23 пары хромосом. Каждой паре хромосом исследователи присвоили определенный порядковый номер. Если у человека хромосом 21-й пары становится три (вместо двух), у него проявляется болезнь Дауна. У больных снижается умственное развитие, продолжительность жизни у них небольшая (обычно не больше 30 лет), размеры головы уменьшены, лицо плоское, разрез глаз косой и т. п. Несмотря на эти недостатки, такие дети отличаются доброжелательностью и послушанием.
VII. Клетки прокариот не имеют сформированного ядра. Их наследственный материал не отделен от цитоплазмы мембранной оболочкой и представлен кольцевой молекулой ДНК, не связанной с ядерными белками. Типичных хромосом, которые в клетках эукариот расположены в ядре, у прокариот нет (однако сам термин «хромосома» часто применяют ДНК прокариот, но в другом значении). Участок цитоплазмы, где расположен наследственный материал прокариот, имеет название ядерная зона, или нуклеоид. Нуклеоид содержит одну молекулу ДНК (чаще всего – замкнутую в кольцо, но встречаются и виды с незамкнутой ДНК) либо несколько ее копий. В цитоплазме клеток многих бактерий наряду с нуклеоидом присутствуют дополнительные кольцевые молекулы ДНК – плазмиды. Их еще называют внехромосомными дополнительными факторами наследственности. От набора плазмид зависит способность прокариот приспосабливаться к изменениям окружающей среды. Например, от наличия или отсутствия определенных генов в составе плазмид зависит устойчивость к определенным антибиотикам.
Литература:
Балан П.Г. Биология: Учебн. для общеобразоват. учебн. заведений: уровень стандарта, академический уровень / П.Г.Балан, Ю.Г.Верес,В.П.АПолищук; пер. с укр. – К.: Генеза, 2010. С. 79 –108.
ТЕМА 6. ЦИТОПЛАЗМА КЛЕТКИ, ЕЁ КОМПОНЕНТЫ
План
I. Одномембранные органоиды.
II. Двумембранные органоиды.
III. Немембранные органоиды.
IV. Клеточные включения.
I. Как ясно из названия к одномембранным относят органоиды, окруженные одной мембраной. Рассмотрим некоторые из них:
1. Эндоплазматическая сеть, или Эндоплазматический ретикулум (сокращенно ЭПС, ЭПР или просто ЭР) – один из важнейших органоидов, присутствующий во всех эукариотических клетках. Представляет собой систему пустот в виде микроскопических канальцев и их утолщений (цистерн). Диаметр канальцев составляет 50–100 нм, а цистерн – до 1000 нм и более. Они ограничены мембраной и соединяются между собой, а также непосредственно переходят в межмембранное пространство оболочки ядра. Площадь мембран ЭПС составляет более половины общей площади всех мембран клетки.
Различают две разновидности эндоплазматической сети: зернистую, или шероховатую, гранулярную и незернистую (гладкую, агранулярную). Зернистая ЭПС получила свое название потому, что на ее мембранах расположены мелкие немембранные органоиды – рибосомы. На рибосомах синтезируется белок, далее его молекулы поступают внутрь цистерн ЭПС, где могут претерпевать дальнейшие модификации (изменять конформацию, присоединять фосфорные, углеводные компоненты и др.) и транспортироваться в другие часть клетки. На мембранах незернистой ЭПС синтезируются липиды, углеводы, определенные гормоны, которые могут накапливаться в ее полостях. В этих полостях (например, в клетках печени) накапливаются и обезвреживаются некоторые токсины. Кроме того, полости ЭПС мышечных клеток, служат внутриклеточным депо (местом хранения) ионов кальция. Это важно для функционирования мышечных волокон: во время их сокращения концентрация ионов кальция резко возрастает, а во время расслабления снижается.
Таким образом, функции ЭПС следующие:
- синтетическая (на мембранах зернистой ЭПС синтезируются белки, гладкой – углеводы и липиды, внутри полостей ЭПС – модификация этих веществ);
- транспортная – транспорт синтезированных веществ внутри клетки, а у растений – и между клетками по плазмодесмам, в которые проникают каналы ЭПС;
- депо ионов кальция
- мембранообразующая – ЭПС служит изначальным источником мембран других органоидов и плазмолеммы (подробнее см. ниже);
-  депо ионов кальция.
депо ионов кальция.
2. Комплекс (или аппарат) Гольджи.Назван в честь итальянского ученого К. Гольджи, который открыл эту структуру. Является обязательным компонентом эукариотических клеток, как и ЭПС, и имеет с последней тесную функциональную связь. Его основной структурной единицей служит диктиосома – скопление одномембранных плоских цистерн, которые не соприкасаются друг с другом, а расположены параллельно на расстоянии около 10 нм. Периферическая часть каждой цистерны часто имеет вид сети канальцев с раздутыми концами. В различных клетках может присутствовать одна или несколько диктиосом. Совокупность всех диктиосом клетки и есть комплекс Гольджи.
Диктиосома обычно располагается в клетке так, что ее сторона, называемая формирующейся, параллельна канальцу ЭПС, от которого время от времени отпочковываются мембранные пузырьки, называемые переходными пузырьками, и присоединяются к крайней цистерне диктиосомы. С противоположной стороны диктиосомы от крайней цистерны (либо от сетчатых краев предыдущих) отделаются пузырьки уже другого качества, называемые пузырьками Гольджи. Относительно механизма перемещения мембран и заключенных в них веществ между разными цистернами диктиосомы существует две гипотезы:
1) с помощью неких промежуточных пузырьков по аналогии с транспортом от ЭПС к диктиосоме;
2) сама диктиосома является динамической структурой: на одном ее конце из переходных пузырьков формируется цистерна, которая постепенно продвигается к другому концу, где распадается на пузырьки Гольджи.
Далее пузырьки Гольджи транспортируются через гиалоплазму и могут сливаться с плазмодеммой, выделяя содержимое во внешнюю среду, либо учувствовать в формировании других одномембранных органоидов (лизосом, вакуоли растительной клетки и др.).
Функции комплекса Гольджи:
- преобразование клеточных мембран:мембрана ЭПС превращается в плазмолемму или мембрану других органоидов, что сопряжено с ее биохимической модификацией, сопровождается изменением толщины и физико-механических свойств; таким образом, все эти мембраны представляют собой единую динамическую систему – концепция эндомембран.
- хранение, модификация и транспорт, выделение из клеткиразличных химических веществ (белков. липидов, угливодов), синтезированных в ЭПС; в частности, ферменты, выделяемые пищеварительными железами животных и человека проходят через аппарат Гольджи.
- Синтез некоторых соединений непосредственно в цистернах диктиосом, например полисахаридов пектинов и гемицеллюлозы, которые затем в пузырьках Гольджи транспортируются к стенке растительной клетки.

3. Лизосомы –органеллы в виде пузырьков диаметром 100–180 нм, окруженных мембраной. Они содержат ферменты, способные расщеплять различные соединения (белки, углеводы, липиды и т. п.). Эти ферменты синтезируются на мембранах зернистой ЭПС, затем проходят через комплекс Гольджи. Из пузырьков Гольджи и формируются лизосомы. В клетке могут быть разныевиды лизосом, отличающихся особенностями строения и функциями. Одни из лизосом, сливаясь с пиноцитозными или фагоцитозными пузырьками, участвуют в формировании пищеварительных вакуолей. При этом активируются ферменты и содержимое вакуоли переваривается. Таким образом, лизосомы обеспечивают процессы внутриклеточного пищеварения. Лизосомы другого типа участвуют в переваривании отдельных частей клетки или всей клетки. Считают, что эти лизосомы уничтожают дефектные органеллы, поврежденные или мертвые клетки, разрушают личиночные органы насекомых и земноводных (например, хвосты и жабры головастиков бесхвостых амфибий) и т. п. Лизосомы могут приближаться к плазматической мембране и выводить свое содержимое – ферменты – наружу. Например, у гриба нейроспоры таким образом обеспечиваются процессы внеклеточного пищеварения.
4. Вакуоли – собирательное название органеллы различных по природе и функциям органоидов в виде пустот, окруженных мембраной и заполненных жидкостью.
Об образовании пищеварительных вакуолей, в которых перевариваются поступающие в клетку соединения и микроорганизмы, мы уже упоминали.
Вакуоли растительных клеток образуются из пузырьков, отделившихся от эндоплазматической сети и комплекса Гольджи. Со временем мелкие вакуоли сливаются в большие, которые могут занимать почти весь объем цитоплазмы. Они заполнены клеточным соком – водным раствором органических и неорганических соединений. Вакуоли поддерживают определенный уровень внутриклеточного давления (тургора), обеспечивая сохранение формы клеток, содержат запасные питательные вещества, конечные продукты обмена или пигменты. Красные, синие, желтые и др. пигменты, растворенные в клеточном соке, определяют окраску отдельных клеток и частей растений в целом (например, плодов вишни, корнеплодов редиса, лепестков цветов и т. п.). Через мембраны вакуолей вещества перемещаются из гиалоплазмы в их полости и наоборот. Благодаря тургорному давлению вакуоли происходит также рост клетки путем растяжения.
В клетках пресноводных одноклеточных животных и водорослей есть сократительные вакуоли – покрытые мембраной пузырьки, способные изменять объем, выводя свое содержимое наружу. Это связано с тем, что в пресной воде концентрация солей значительно ниже, чем в цитоплазме. Поэтому согласно физическим законам вода из окружающей среды поступает в клетку, повышая давление внутри нее. Излишек жидкости накапливается в сократительной вакуоле. Стенка вакуоли сжимается благодаря сократительным белкам, входящим в ее состав, и выталкивает жидкость наружу из клетки. Таким образом, сократительные вакуоли регулируют внутриклеточное давление, выводя излишки воды из клетки и предотвращая ее разрушение. Кроме того, эти органеллы обеспечивают выведение некоторых растворимых продуктов обмена веществ. Деятельность сократительной вакуоли имеет значение и для газообмена, поскольку способствует поступлению в клетку воды с растворенным кислородом и выведению углекислого газа. У инфузории строение сократительных вакуолей наиболее сложное. Например, у инфузории-туфельки вакуоль состоит из сократительного пузырька (резервуара), который открывается наружу отверстием – порой. В пузырек впадают длинные тонкие радиальные каналы, в которые из цитоплазмы поступают водные растворы. Сначала сокращаются стенки канальцев, проталкивая жидкость к резервуару, а затем сокращается наполненный резервуар, выводя содержимое через пору.
5. Пероксисомы – округлые одномембранные органеллы диаметром около 0,3–3,5 мкм. Пероксисомы обнаружены у различных ор ганизмов: одноклеточных и многоклеточных животных, дрожжей, высших растений, водорослей. Эти органеллы часто расположены вблизи мембран эндоплазматической сети, митохондрий, пластид. В них содержатся разнообразные ферменты, в частности те, что обеспечивают преобразование жиров в углеводы или способны расщеплять перекись водорода Н2О2 на кислород и воду. У растений пероксисомы участвуют в процессах поглощения кислорода во время световой фазы фотосинтеза.

II. Двумембранные (двухмембранные) органоиды– органеллы клеток эукариот, поверхностный аппарат которых состоит из двух мембран, разделенных межмембранным промежутком. Они пространственно не связаны с другими органеллами и принимают участие в энергетическом обмене. Их всего два вида: митохондрии и пластиды. Иногда к двумембранным органоидам причисляют также ядро на основании того факта, что ядерная оболочка тоже состоит из двух мембран. Однако сходство это чисто внешнее, с митохондриями и пластидами по своей природе и происхождению ядро не имеет ничего общего.
1. Митохондрии – органеллы клеток большинства видов растений, грибов и животных. Их нет лишь у некоторых одноклеточных эукариот, которые обитают в бескислородной среде, – анаэробов. Митохондрии служат своеобразными клеточными «генераторами энергии». Они имеют вид сфер, палочек, иногда разветвленных нитей (длиной 0,5–10 мкм и более). Число этих органелл в клетках разных типов может варьировать от 1 до 100 000 и более. Оно зависит от того, насколько активно происходят процессы обмена веществ и преобразование энергии. Так, клетка значительных размеров амебы хаос содержит до 500 000 митохондрий, тогда как в мелкой клетке паразитических жгутиконосцев – трипаносом (возбудителей сонной болезни человека) есть лишь одна гигантская разветвленная митохондрия.
Наружная мембрана митохондрии гладкая, а внутренняя образует складки, направленные внутрь органеллы – кристы. Кристы имеют вид дискообразных, трубчатых или пластинчатых, зачастую разветвленных, образований. На поверхности крист, граничащей с внутренней средой митохондрии, есть особые грибовидные белковые образования – АТФ-сомы (от греч. сома – тело). Они содержат комплекс ферментов, необходимых для синтеза АТФ. Внутреннее пространство митохондрий заполнено полужидким веществом – матриксом. Там находятся рибосомы, молекулы ДНК, иРНК, тРНК и др. и синтезируются белки, входящие в состав внутренней мембраны.
Основная функция митохондрий – синтез АТФ. Этот процесс происходит за счет энергии, которая высвобождается во время окисления органических соединений. Начальные реакции происходят в матриксе, а завершающие – на внутренней мембране митохондрий.
2. Пластиды – органоиды клеток растений и некоторых одноклеточных животных (например, эвглены зеленой). Известно три типа пластид – хлоропласты, хромопласты и лейкопласты, различающиеся по окраске, особенностям строения и функциям.
Хлоропласты – пластиды, обычно окрашенные в зеленый цвет благодаря наличию пигмента хлорофилла. Но в клетках многих групп водорослей (красных, бурых и т. п.) их цвет может быть иным. Это объясняется тем, что в них, кроме хлорофилла, есть и другие пигменты – красные, желтые, бурые и др.
Как и у митохондрий, наружная мембрана хлоропластов гладкая, а внутренняя образует выросты, направленные внутрь. Строма – комплекс веществ, заполняющих внутреннее пространство хлоропласта. С внутренней мембраной связаны особые структуры – тилакоиды. Это плоские одномембранные цистерны. Большие тилакоиды расположены одиночно, а более мелкие собраны в граны (группы по 5–20 шт., напоминающие стопки монет). В тилакоидах содержатся основные (хлорофиллы) и вспомогательные (каротиноиды) пигменты, а также все ферменты, необходимые для осуществления фотосинтеза. В строме хлоропластов есть молекулы ДНК, разные типы РНК, рибосомы, зерна запасного полисахарида (преимущественно крахмала). Основная функция хлоропластов – осуществление фотосинтеза. Кроме того, в них, как и в митохондриях, на мембране тилакоидов имеются АТФ-сомы и происходит синтез АТФ. Также в хлоропластах синтезируются некоторые липиды, белки мембран тилакоидов, ферменты, обеспечивающие реакции фотосинтеза.
Лейкопласты – бесцветные пластиды разнообразной формы, в которых запасаются некоторые соединения (крахмал, белки и т. п.). В отличие от хлоропластов, у лейкопластов внутренняя мембрана может образовывать лишь немногочисленные тилакоиды. В строме лейкопластов содержатся рибосомы, ДНК, разные типы РНК, ферменты, обеспечивающие синтез и расщепление запасных веществ (крахмала, белков и т. п.). Лейкопласты могут быть полностью заполнены зернами крахмала.
Хромопласты – пластиды, окрашенные в разные цвета (например, желтый, красный, фиолетовый). Цвет этим пластидам придают различные пигменты (преимущественно каротиноиды), которые в них накапливаются. Поскольку хлорофилл в хромопластах отсутствует, то зеленой окраски они не имеют. Хромопласты обесцвечивают расцветку лепестков цветов, плодов, листьев и других частей растений. Внутренняя система мембран в хромопластах отсутствует или же образована отдельными тилакоидами.
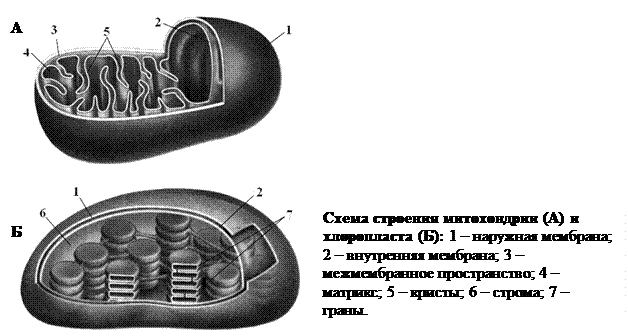 Все специализированные пластиды: хлоропласты, хромопласты и лейкопласты – могут развиваться из мелких бесцветных пердшественников – пропластид – содержащихся в клетках зародыша семени либо в меристемах (образовательных татканях) взрослого растения. Возможен также обратный процесс – упрощение структуры специализированных пластид и превращение их снова в пропластиды. Помимо этого разные типы специализированных пластид при определенных условиях могут превращаться друг в друга. Так, превращение хлоропластов в хромопласты можно наблюдать при созревании плодов и пожелтении листьев осенью. При этом в них разрушается хлорофилл и накапливаются каратиноиды. Лейкопласты могут превращаться в хлоропласты, например, в клубнях картофеля на свету: в них исчезает крахмал, образуются тилакоиды и граны, синтезируется хлорофилл. Образование лейкопластов и хлоропластов из хромопластов наблюдается реже, например, при старении некоторых окрашенных тканей отдельных видов растений.
Все специализированные пластиды: хлоропласты, хромопласты и лейкопласты – могут развиваться из мелких бесцветных пердшественников – пропластид – содержащихся в клетках зародыша семени либо в меристемах (образовательных татканях) взрослого растения. Возможен также обратный процесс – упрощение структуры специализированных пластид и превращение их снова в пропластиды. Помимо этого разные типы специализированных пластид при определенных условиях могут превращаться друг в друга. Так, превращение хлоропластов в хромопласты можно наблюдать при созревании плодов и пожелтении листьев осенью. При этом в них разрушается хлорофилл и накапливаются каратиноиды. Лейкопласты могут превращаться в хлоропласты, например, в клубнях картофеля на свету: в них исчезает крахмал, образуются тилакоиды и граны, синтезируется хлорофилл. Образование лейкопластов и хлоропластов из хромопластов наблюдается реже, например, при старении некоторых окрашенных тканей отдельных видов растений.
Хлоропласты и митохондрии, в отличие от других органелл, характеризуются определенной степенью автономии (независимости от других частей клетки):
- содержат собственную наследственную информацию – кольцевую молекулу ДНК, напоминающую ДНК из ядерной зоны клеток прокариот;
- имеют аппарат, который осуществляет синтез собственных белков (рибосомы, а также все виды РНК);
- в отличие от других органелл, не возникают из других мембранных структур клетки, а размножаются делением.
Поэтому большинство ученых предполагает, что митахондрии и пластиды являются потомками прокариотических организмов – внутиклеточных симбионтов эукариот. Но следует отметить, что их автономия не абсолютна: в частности часть ферментов необходимых для жизнедеятельности митахондрий и пластид синтезируется е в них самих, а в ядре клетки. Последовательности ДНК митохондрий и пластид, кодировавшие эти белки, были утеряны в ходе эволюции.
III. Группа немембранных органоидов объединяет органоиды с различно природы, характеризующиеся одной общей чертой – отсутствием мембран в своей структуре.
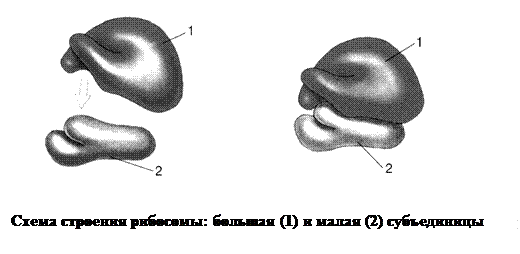
1. Рибосомы – мельчайшие (22х17 – 26х19 нм) лишенные поверхностной мембраны органеллы, участвующие в синтезе белков. Они присутствуют в клетках как прокариот, так и эукариот, имеют вид сферических телец, состоящих из двух разных по размерам частей – субъединиц: большой и малой. Каждая из субъединиц состоит из взаимодействующих между собой рРНК и белков. Субъединицы рибосом могут разъединяться после завершения синтеза белковой молекулы и вновь соединяться между собой перед его началом. Субъединицы образуются в ядрышке: на молекуле ДНК синтезируется рРНК, которая соединяется с особыми рибосомальными белками, поступающими из цитоплазмы. Готовые субъединицы транспортируются в цитоплазму. Рибосомы митохондрий и пластид меньше расположенных цитоплазме по размерам, но подобны по строению. Количество рибосом в клетке зависит от интенсивности процессов биосинтеза белков.
2. Микронити, или микрофиламенты, – тонкие нитевидные структуры, диаметром 4–7 нм, которые состоят из сократительных белков, преимущественно актина. Они пронизывают цитоплазму и могут образовывать сплетение под плазматической мембраной. Пучки микронитей прикреплены одним концом к определенной структуре (например, плазматической мембране), а вторым – к другой (органелле и т.п.). Микрофиламенты участвуют в изменении формы клетки, например при амебоидном движении, процессах поступления в клетку и выводе из нее различных соединений, перемещении органелл внутри клетки т. п. При делении некоторых клеток животных они образуют особое сократительное кольцо, разделяющее цитоплазмы дочерних клеток.
3. Микротрубочки – полые цилиндрические структуры диаметром 10–25 нм, стенки которых состоят из спирально уложенных глобул белка белком тубулина. Они могут собираться и разъединяться, соответственно микротрубочка удлиняется или укорачивается. микротрубочки участвуют в формировании веретена деления эукариотических клеток, входят в состав ресничек, жгутиков, центриолей. Микротрубочки обеспечивают перемещение органелл и макромолекул в клетке. При этом пучки микротрубочек одним концом прикрепляются к одной структуре или молекуле, а вторым – к другой.
Микротрубочки и микрофиломенты благодаря достаточно жесткой структуре выполняют еще одну важную роль в клетке – образуют цитоскилет.

4. Клеточный центр – органелла, которая состоит из двух центриолей, расположенных в светлом уплотненном участке цитоплазмы. Центриоли имеют вид пустого цилиндра, состоящего из девяти комплексов микротрубочек, по три в каждом. В периоды между делениями центриоли расположены вблизи ядра, обычно рядом с комплексом Гольджи. Во время деления клетки центриоли расходятся к ее полюсам и служат центрами организации нитей веретена деления. Поэтому каждая центриоль попадает в одну из дочерних клеток. Центриоли удваиваются в период между делениями, при этом новая образуется путем самосборки. Кроме того, центриоли участвуют в формировании микротрубочек цитоплазмы, жгутиков и ресничек. В клетках, где центриоли отсутствуют (например у высших растений), процессы формирования микротрубочек веретена деления и органелл движения происходят и без их участия.

5. Жгутики и реснички имеются у многих одноклеточных организмов (хламидомонада, вольвокс, эвглена, инфузории), а также характерны для некоторых клеток многоклеточных (клетки мерцательного эпителия дыхательных путей млекопитающих, сперматозоиды животных, высших споровых растений и т. п.). Жгутики и реснички имеют вид тонких выростов цитоплазмы, покрытых плазмолеммой. Внутри этих органелл расположена сложная система микротрубочек – аксонема. На поперечном срезе через внеклеточную часть жгутика или реснички заметно, что по периферии расположено кольцо из девяти комплексов микротрубочек, по две в каждом, и еще две микротрубочки – в центре. Центральные микротрубочки выполняют опорную функцию, а периферические – обеспечивают движение. Часть жгутиков и ресничек, расположенная в цитоплазме, имеет название базального тельца. Оно состоит из девяти периферических комплексов по три микротрубочки в каждом.
Жгутики и реснички двигаются за счет высвобождения энергии АТФ, при этом соседние пары микротрубочек как будто скользят одна относительно другой. Хотя строение жгутиков и ресничек похоже, характер их работы разный. Работа ресничек в общем напоминает греблю веслами и обычно скоординирована (например, у инфузорий). Для жгутиков характерно вращение или волнообразное движение. Работа жгутиков и ресничек обеспечивает не только передвижение, но и поступление частичек пищи к поверхности клетки вместе с потоками воды, которые они создают (например, движение жгутиков пищеварительных клеток гидры). Жгутики и реснички могут также выполнять чувствительную (например, у ресничных червей) и защитную (реснички эпителия носовой полости) функции.
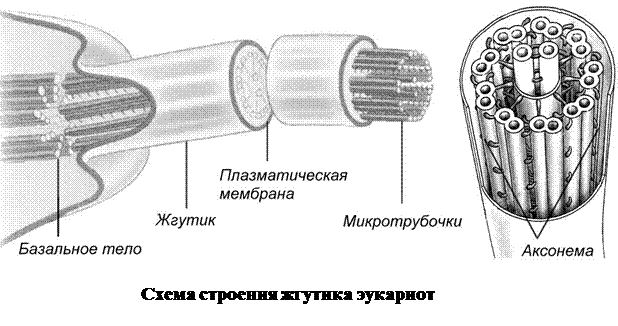 Базальные тельца жгутиков и ресничек по строению напоминают центриоли клеточного центра.
Базальные тельца жгутиков и ресничек по строению напоминают центриоли клеточного центра.

IV. Клеточные включения,в отличие от органелл, являются непостоянными компонентами клетки. Они могут появляться и исчезать в процессе ее жизнедеятельности. Обычно это запасные вещества. Клеточные включения содержатся в цитоплазме, вакуолях или пластидах растительных клеток в растворенном (капли жиров) или твердом (кристаллы, волоконца, зерна и т. п.) состояниях. В клетках грибов, животных и человека запасается полисахарид гликоген (в виде зерен или волоконец), различные липиды и белки (например, «желток» яйцеклеток). В цитоплазме могут накапливаться и нерастворимые продукты обмена: соли мочевой кислоты, кристаллы щавелевокислого кальция (в щавле, бегонии и др.). Под действием ферментов большинство клеточных включений распадается на соединения, участвующие в обмене веществ. Они могут использоваться клеткой в процессе роста, растениями – во время цветения, созревания плодов и т. п.
У некоторых одноклеточных животных есть особые внутриклеточные структуры, выполняющие опорную функцию. Как и включения, это конструкции определенной формы, расположенные в гиалоплазме и не ограниченные мембранами. Например, в цитозоле паразитических одноклеточных животных лямблий есть опорный стержень, состоящий из органического вещества. У морских одноклеточных животных радиолярий капсула из органического вещества разделяет цитоплазму на внешнюю и внутреннюю части, свойства которых различны. Кроме того, эти животные имеют сложно устроенный внутриклеточный скелет в виде вложенных один в другой дырчатых слоев, корон и т. п., состоящий из SiО2 или SrSO4.

Литература:
Балан П.Г. Биология: Учебн. для общеобразоват. учебн. заведений: уровень стандарта, академический уровень / П.Г.Балан, Ю.Г.Верес,В.П.АПолищук; пер. с укр. – К.: Генеза, 2010. С. 112 –130.
| <== предыдущая лекция | | | следующая лекция ==> |
| ТЕМА 4. ОРГАНИЧЕСКИЕ ВЕЩЕСТВА ЖИВЫХ ОРГАНИЗМОВ, ИХ РАЗНООБРАЗИЕ И БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ | | | Раздел: Гнойная хирургия |
Дата добавления: 2014-12-15; просмотров: 5564;