ЭЛЕКТРОШОК И ПСИХИЧЕСКОЕ СОСТОЯНИЕ 5 страница
ТЕОРИИ ЭМОЦИИ
выраженные изменения приводят к состоянию депрессии. Сдвиг равновесия в сторону преобладания симпатической системы приводит к бодрствованию и ощущению счастья, тогда как у животных проявляется только усиление агрессивных реакций. Повышение психических функций воспринимается человеком как ощущение успеха. Усиление типоталамо-кортикальных разрядов у животных может быть связано с усилением агрессивности, а у человека с повышением -психомоторной активности и маниакальным поведением.
Кофеин и фенамин ускоряют и усиливают условные реакции [246] и замедляют угашение условных рефлексов, в то время как депресоирующие агенты, такие, как алкоголь и амитал натрия, оказывают противоположное действие.
Эти наблюдения поддерживают гипотезу о том, что депрессия связана с преобладанием трофотропной системы, а подъем—с преобладанием эрготротшой. Ай-зенк [230] считает, что эти состояния связаны с определенными особенностями личности (интроверзия и экстро-верзия), а также с невротическими явлениями.
Нам кажется вероятным, что 'индивидуальные различия реакций на зти препараты зависят от характера равновесия систем .гипоталамуса [919], однако, чтобы доказать правильность этого предположения, необходимы дальнейшие эксперименты.
Значение этих выводов для нейропсихиатрии очевидно. Можно предполагать, что некоторые депрессивные состояния связаны с повышением реактивности центральных парасимпатических структур и снижением активности симпатических механизмов. Вторую часть этого предположения подтверждает проба с мехолилом, однако экспериментальных данных, подтверждающих первую его часть, пока не существует. При состояниях, связанных с агрессивностью и ненавистью, предполагаемое повышение реактивности симпатической системы действительно подтверждается пробой с мехолилом и секрецией норадреналина. С нашей концепцией согласуются также данные о повышении секреции норадренаг •пина при маниакальном состоянии и о ее понижении при Депрессии.
S34
ГЛАВА XXV
Основы качественных различий эмоций
Теорию, которая не раскрывает механизмы, определяющие количественные и качественные различия эмоций, нельзя считать завершенной как в теоретическом, так и в практическом плане. Поэтому мы рассмотрим прежде всего физиологические основы качественных различий эмоций.
Мы строим наш анализ не на центральных процессах, лежащих в основе эмоций, а на сопровождающих их периферических разрядах в вегетативной нервной системе. Еще в 1924 г. Олпорт [17] связывал приятные и неприятные эмоции соответственно с парасимпатической и симпатической системами. Действительно, точно установлено, что вид и запах пищи возбуждают парасимпатические влияния на желудочно-кишечную систему и ее железы и что в состоянии пищевого удовлетворения активность эрготропной системы снижается, а парасимпатической— повышается. Однако не нужно идти далеко, чтобы обнаружить вопиющие противоречия. Печаль сопровождается плачем, возникающим в результате парасимпатической импульсации, а возмущение связано с усилением перистальтики, расширением сосудов слизистых оболочек и прочими симптомами. Не только приятный запак съедобной пищи, но и неприятные запахи стимулируют секрецию, регулируемую парасимпатическими нервами, и могут вызвать рвоту. Вместе с тем усиленная перистальтика мочевого пузыря и прямой кишки, характерные симптомы возбуждения парасимпатической системы сопровождают такие ощущения, как страх и беспокойство.
Со времени работ Кэннона общепризнано, чго испуг стимулирует симпатические или симпато-адреналовые разряды или, по терминологии Гесса, вызывает активацию эрготропной системы. Однако примеры, приведенные здесь и в гл. IV, показывают, что это утверждение в некоторых случаях является чересчур узким, а в других— совсем неверным. Так, тяжелые эмоциональные расстройства связаны и с симпатическими, и с парасимпатическими разрядами, первые из которых обнаруживаются по реакциям сосудов, а вторые — по изменениям
ТЕОРИИ ЭМОЦИИ 535
со стороны внутренних органов (иногда половых). В качестве одного из примеров патологической настройки гипоталамуса [310] укажем, что при неврозах эмоциональный стресс вызывает падение уровня сахара крови (через ваго-инеулиновую систему), а не его повышение [12, 722 а].
Если ограничить наше обсуждение нормальными реакциями, то следует признать, что эмоции умеренной интенсивности, если они приятны, связаны преимущественно с парасимпатическими разрядами, а если они неприятны и болезненны, то с симпатическими. При повышении степени эмоционального возбуждения нисходящие гипоталамические разряды не ограничиваются одним отделом вегетативной нервной системы. Наиболее выразительным примером приятных эмоций высокой интенсивности, вызывающих такой тип разрядов, является оргазм, при .котором сосудистые и соматические изменен.ия указывают на активацию эрготропной системы, а реакция половых органов обнаруживает одновременные парасимпатические разряды. Такое двойное действие отражает двойственную природу процесса: он занимает пограничное место между удовольствием и болью'. Однако в нормальном организме тяжелые (неприятные) эмоции также включают симпато-адреналовую и вато-инсу-линовую системы при преобладании первой. В состояниях предельного испуга и ужаса циркуляторный коллапс и замедление сердечного ритма являются результатом возросшей парасимпатической активности, а расширение зрачка, потоотделение свидетельствуют о повышении симпатических разрядов. В этом случае наблюдается также диссоциация между симпатической активностью и тонусом скелетной мускулатуры2.
Наиболее значительным выводом из этих рассуждений является представление о сложной зависимости эмо-
* * * *
1 Хордотомия (перерезка спино-таламического тракта в передне-латеральном пучке .спинного мозга) вызывает исчезновение оргазма, хотя рефлекторно вызываемая эрекция и эйякуляция сохраняются.
' Напомним также о возникновении тошноты и рвоты при условном сигнале болевого раздражителя у кошки, близкой к невротическому состоянию (си. гл. XVI).
536 ГЛАВА XXV
ций от вегетативной нервной системы, особенно при высокой интенсивности. Хотя в организме, находящемся в состоянии покоя, сдвиг >в равновесии вегетативных систем на уровне гипоталамуса определяется принципам реципрокной иннервация [356], при сильном эмоциональном возбуждении (а также при других условиях, вызывающих интенсивное возбуждение вегетативных центров, например при судорогах, вызванных электрошоком [239, 529]) происходит отклонение от этого правила.
Поскольку изменения вегетативного равновесия в гипоталамусе, вызываемые в эксперименте у животного раздражением, введением фармакологического препара-та или разрушением, приводят к изменениям восходящих и нисходящих разрядов гипоталамуса [59, 310, 549] и поскольку первые связаны с соответствующими «психическими» сдвигами, возникает вопрос о том, каковы характерные отношения, существующие между нисходящими вегетативными разрядами, с одной стороны, и качеством и интенсивностью эмоций — с другой.
Для подтверждения наличия специфических отношений между «неприятными» эмоциями и симпатическими разрядами напомним читателю эксперименты по после дованию ярости: 1) при естественных условиях; 2) в результате раздражения симпатического отдела гипоталамуса различными методами; 3) после высвобождения активности этого отдела путем устранения тормозных влияний со стороны вентро-медиального ядра гипоталамуса [943]. Сходные результаты были получены при разрушениях перегородки [94]. Напротив, отношения между парасимпатической системой и приятными эмоциями, очевидно, выступают в опытах Гесса с раздражением перегородки и переднего отдела гипоталамуса [641]; в этих опытах облизывание и другие поведенческие реакции позволяли предположить возникновение у животных ощущения удовольствия. Подобный эффект удалось получить также с различных частей висцерального мозга, например с гиппокампа (особенно при химическом раздражении [603]). Более того, раздражение гиппокампа сначала сопровождалось периодом разрядов последействия, связанных с признаками возбуждения симпатического отдела (расширение зрачка, пилоэрекция), а
ТЕОРИИ ЭМОЦИИ
затем фазой «благополучия», характеризующейся вылизыванием и другими проявлениями материнского инстинкта по отношению к котенку, к которому до этого животное проявляло агрессивную реакцию [698]. В этом случае вновь сдвиг вегетативного равновесия сопутствует фундаментальным изменениям эмоционального поведения. Тесная связь удовольствия и половых реакций со сниженной реактивностью к болевым раздражителям часто отмечалась в экспериментах с раздражением лим-бической системы [18].
В то время как раздражение миндалевидного ядра вызывает изменения поведения, указывающие на появление страха и ярости [959], удаление миндалевидного комплекса у кошек приводит к гиперсексуальности .и к «усилению .реакций удовольствия на поглаживание и похлопывание» [541]. При этом эмоциональные реакции, связанные с усилением разрядов в симпатическом отделе гипоталамуса, такие, как страж и агрессивное поведение, а также реактивность к болевым раздражителям, оказываются сниженными. Однако эти характерные сдвиги не 'возникают, если перед амигдалэктомией произвести двухстороннее разрушение вентро-медиальных ядер гипоталамуса, или устраняются, если эти ядра' разрушить после амигдалэктомии. Такие животные обнаруживают поведение, характеризующееся одичанием, чго обычно наблюдается у кошек с указанными разрушениями гипоталамуса. Отсюда следует, что амигдалэктомия приводит к повышению активности вентро-медиальных ядер, которые, как известно, тормозят активность симпатического отдела гииоталагмуса. Эти эксперименты показывают, что разрушения висцерального мозга изменяют эмоциональное поведение через гипоталамус1.
1 Эксперименты Всйекранца и Вильсона [936] в соответствии с описанной выше работой показали, что после амигдалэктомии у обезьян наблюдается повышение порога реакции избегания, вызываемой болевым раздражителем. Оперированные животные по-прежнему реагируют па резерпин, подобно нормальным. Оба эти факта можно объяснить на основании данных, приведенных здесь и в гл. III и показывающих, что лимбическая система изменяет возбудимость гипоталамуса и вызывает сдвиг в равновесии его систем и что резерпин действует на промежуточный мозг.
ГЛАВА XXV
Однако «е следует забывать, что экспериментальные даиные по раздражению и разрушению лимбичеокой системы довольно противоречивы. В какой мере несоответствие зависит от видовых различий [40, 545] и локализации разрушения [959], .не вполне ясно. Лабильность этих функций .позволила Глуру [382] предположить, что «миндалевидное ядро ,и другие части обонятельного мозга модулируют активность, интегрированную в подкорковых структурах». Поскольку активность лимбичеокой системы тесно связана с гипоталамусом, если не зависит от него полностью, следует помнить, что уровень реактивности гипоталамуса весьма изменчив и что равновесие его систем варьирует в широких пределах [310].
В пользу предположения о том, что раздражение или повреждение некоторых частей лимбической системы изменяет эмоциональное поведение через сдвиг равновесия систем .гипоталамуса, свидетельствуют эксперименты на крысах, у которых последовательно разрушали перегородку и миндалевидное ядро. Мы уже видели, что раздражение перегородки вызывает положительные реакции н тормозит эртотропную систему, а раздражение миндалевидного ядра вызывает страх и ярость [289]. Наоборот, разрушение миндалевидного ядра снижает эмоциональные реакции (это особенно отчетливо видно на неприрученных, диких крысах, которые становятся ручными после операции [961]), а разрушения перегородки приводят к резкому их повышению [537]. Этот эффект полностью устраняется последующим удалением миндалевидного ядра; в контрольной группе, у животных с разрушениями в области новой коры и поясной извилины, обнаруживается лишь незначительное снижение эмоциональной реактивности. Далее, повышение эмоциональных реакций после разрушений перегородки было значительно менее выражено у крыс с ам.итдалэкто-мией, чем у животных контрольной оперированной группы (фиг. 53).
Если правильно наше утверждение о связи сдвигов в равновесии вегетативных центров гипоталамуса со сдвигами эмоционального состояния, то можно ожидать, что патологические процессы у человека, приводящие к усилению симпатических или парасимпатических разря-
ТЕОРИИ ЭМОЦИИ
дов, должны сопровождаться различными эмоциональными состояниями. Поэтому имеющиеся у «ас сведения об эмоциях и их связи с гипоталамусом, которые до сих пор черпались главным образом из наблюдений над
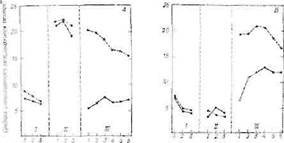
Фиг. 53. Интенсивность эмоциональных реакций у крыс после разрушения перегородки, миндалевидного ядра и коры [537].
А. Повышение интенсивности эмоциональных реакций после разрушения перегородки, исчезающее после разрушения миндалевидного ядра; 15 крыс были разделены на 2 группы; установив контрольный уровень эмоциональных реакций (/). в обеих группах произвели разрушение перегородки, в результате чего резко усилились эмоциональные реакции (//). Затем у 10 крыс (сплошная линия) произвели разрушение миндалевидного ядра (///) — уровень их эмоциональных реакций восстановился. У других 5 крыс (пунктирная линия) были произведены разрушения поясной извилины и новой коры для контроля влияния самого оперативного вмешательства; у них обнаружилось лишь незначительное и постепенное снижение эмоциональных реакций.
Б. 1 — контроль; // — отсутствие изменений эмоциональных реакций после разрушений коры (новая кора или поясная извилина) или миндалевидного ядра; /// — крысы с разрушениями перегородки после повреждения коры обнаруживают повышение эмоциональных реакций, сравнимое с эффектом разрушения одной только перегородки, в то время как аналогичные разрушения перегородки после амигдалэктомии вызывают значительно менее выраженное усиление эмоциональных реакций. На оси абсцисс обозначены дни после операции.
эмоциональным поведением животных, должны пополниться за счет словесных отчетов больных. Особый интерес представляют наблюдения над больными с цирку-ляторными нарушениями диэнцефапьного происхождения. Как показано в гл. XV, у таких больных бывают приступы крайнего возбуждения системы блуждающего нерва (замедление ритма или полная остановка сердца и падение кровяного давления) или симпатической системы (максимальное ускорение ритма и повышение
ГЛАВА XXV
кровяного давления). Поведение этих двух групп больных существенно различается [112]. Больные с сосудистыми приступами парасимпатического происхождения обычно тихие, довольно спокойные, они жалуются на слабость, 'головокружение и приступы in отер и сознания, но не производят 'впечатление людей, взволнованных своим состоянием. Больные, страдающие от приступов симпатического происхождения, наоборот, выражают крайнее беспокойство, сильно напуганы (Vernichtungs-gefiihl), хотя объективные данные указывают на относительную безопасность их состояния. Для первой группы характерно состояние, близкое к расслаблению и сну, для второй — состояние максимального эмоционального возбуждения, хотя у больных первой труппы значительно больше оснований для беспокойства. Связь между парасимпатическими разрядами .в системе гипоталамуса и покоем, с одной стороны, и между симпатическими разрядами и эмоциональным возбуждением, с другой, подтверждается в экспериментах, где подобные сдвиги активности вегетативной системы вызывали у человека рефлекторно путем введения норадреналина и мехолила соответственно [311]. Введение норадреналина приводило к рефлекторному повышению парасимпатической активности и успокоению, а у некоторых больных вызывало сон, тогда как вызванные мехолилом симпа-гичеокие разряды сопровождались ощущением напряжения in раздражительностью. Эти наблюдения подтверждают нашу гипотезу о том, что центральные (гипоталамические) разряды, зависящие, очевидно, от разных частей гипоталамуса, приводят к принципиально различным восходящим и нисходящим влияниям. Предполагается, что характер восходящего разряда, в значительной степени определяющий характер эмоции, зависит не только от того, какая область гипоталамуса возбуждена, но и от обратной связи, идущей от иннервируе-мых вегетативной нервной системой органов. Только в этом смысле можно говорить о том, что периферические вегетативные реакции влияют на состояние гипоталамуса и таким образом модулируют эмоции.
Кроме того, эти рассуждения показывают, что различные эмоции нельзя объяснять только на основании
ТЕОРИИ ЭМОЦИИ 541
фактора интенсивности, как это делает активационная теория. В .настоящее время .кажется, что качественные различия .между эмоциями, которые на основе психологического анализа требуют трехмерного изображения [802], физиологически зависят от парасимпатических, симпатических .и смешанных разрядов, связанных с соответствующими дифференцированными формами активации висцерального мозга и .новой коры. Далее, следует помнить, что эмоциональное возбуждение по крайней .мере при наиболее высоких уровнях ведет к высвобождению яорадреналина, адреналина и инсулина, а также к регулируемой гипотала1мусом секреторной активности гипофиза. Весьма вероятно, что на изменение характера гипоталамо-корт.икальных разрядов влияют не только адреналин и норадреналин, действие которых на ретикуло-шпоталамическую систему было подробно изучено [768], но и другие гормоны, высвобождаемые в результате активности •гипоталамо-гипофизарной системы [960]. С повышением степени возбуждения гипоталамуса возникают изменения качества эмоций, например наблюдается переход от страха ,к ярости [289]. Предполагается, что это изменение представляет собой не только результат .возрастающей частоты разрядов и увеличения числа разряжающихся нейронов, но также следствие изменения конфигурации разрядов, связанных со сдвигами тонуса и активности поперечнополосатых мышц. Обратная связь от соматической нервной системы при страхе и ужасе иная, чем при ярости; роль проприоцептивной импульсации для поддержания возбудимости типотала.муса и интенсивности типотала.мо-кор-тикальных разрядов была показана в гл. XVII. Этим объясняется тот факт, что легкая кураризация может привести .к превращению эмоциональной реакции ярости в реакцию страха вследствие снижения проприоцептивной импульсации в расслабленных мышцах [311].
О значении интенсивности эмоций
До сих пор мы подчеркивали качественные изменения периферических вегетативных разрядов, сопровождающих различные виды эмоционального возбуждения,
ГЛАВА XXV
однако количественный аспект не менее важен. Линдслей [579], а также другие авторы отметили наличие непрерывного перехода от она' к эмоциональному возбуждению, которое выражается сдвигами в электроэнцефалограмме и указывает на возрастание степени активности ретикуло-гипоталамической системы. В то время как но мере перехода от «настороженного внимания» к сильным эмоциям, таким, как страх или ярость, степень активации как бы постепенно нарастает, это постепенное нарастание не распространяется на .все поведенческие, вегетативные и электроэнцефалографические изменения. Так, при эмоциональном возбуждении у человека внимание на определенном объекте (раздражителе) концентрируется с трудом, реактивность к раздражителям симпатической системы снижается (возможно, на гипо-таламическом уровне), а спонтанная активность возрастает (фиг. 54). Эти явления, изучавшиеся у человека в опытах с кожно-тальваническим рефлексом, очевидно,
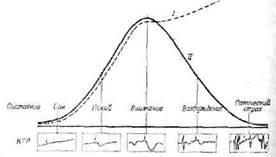
Фиг. 54. Теоретические отношения между степенью активации (/) и реакцией на электрическое раздражение (//) [151].
КГР - кожно-гальваничсский рефлекс; каждая стрелка указывает момент нанесения специфического стандартного раздражения а отклонения соответствуют сдвигу кожного сопротивления. Иа трех первых стадиях активации кожно-гальванический рефлекс возникает в ответ на ««ирую, ;ее Раздражу ние. причем его амплитуда прямо зависит от степени активации Однако в состояниях возбуждения и паники становятся все более частыми «понтаи ны» сдвигиГКГ'Г> (неизвестные раздражители), а интенсивность реакции на стандартный раздражитель снижается.
ТЕОРИИ ЭМОЦИЙ
связаны с усилением спонтанных симпатических разрядов, которые можно обнаружить ло сокращению третьего века у подопытного животного после внутривенного ГЗЗЗ] или интратипоталамичеокого введения метразола [325]. Очевидно, состояние резко повышенного симпатического возбуждения гипоталамуса характеризуется повторными «спонтанными» симпатическими разрядами.
Предполагается, что при «сильных» эмоциях гипоталамус дает почти максимальные разряды лод влиянием разрядов от ретикулярной формации и органов чувств, причем импульсы от последних поступают на ретикулярную формацию и гипоталамус не только непосредственно, но также через новую и димбическую кору. При этом дифференциация процесса активации и 'функций, принадлежащих различным кортикальным областям в нормальных физиологических условиях, снижается [581]. Возникающая вследствие этого «функциональная» декортикация не является результатом «кортикального конфликта», ведущего к высвобождению подкорки, как предполагает Дарроу [163], а зависит от чрезмерного возбуждения системы гипоталамуса, которое несовместимо с дифференцированным действием коры, необходимым для внимания [7] и высших психических процессов.
При дальнейшем усилении возбуждения гипоталамуса возникают невротические явления.
Гипоталамическая система
при экспериментальных неврозах
Так называемым экспериментальным неврозам предшествуют и их сопровождают глубокие изменения эмоционального поведения и вегетативной активности. Бели применяются относительно мягкие средства воздействия и длительное невротическое состояние не развивается, условный раздражитель вызывает сначала типичные симптомы возбуждения симпатической системы, затем возникают отчетливые парасимпатические явления, например рвота [510]. В гл. XVI было показано, что три таких явлениях, как тошнота .и рвота, доминирует пара-
544 ГЛАВА XXV
симпатическая система, а эрготропная система тормозится. Этот сдвиг вегетативного равновесия, сопровождающий тяжелые эмоциональные расстройства, можно рассматривать как а-омеастатичеокую реакцию: снижается реактивность в отношении всех окружающих раздражителей, и особенно тех, которые вызывают серьезные сдвиги.
Если условный раздражитель перестает подкрепляться, рвота -исчезает и восстанавливается нормальное равновесие в вегетативной системе. Вместе с тем, если на некоторое время насильственно подавляется соответствующая реакция на условный пищевой раздражитель или если используются другие способы индукции невроза, то возникают патологические условные рефлексы в форме общего возбуждения или торможения. При этих условиях обычно возникают 'расстройства вегетативных функций; нарушения сердечного ритма могут сохраняться годами [285]. Очевидно, при развитии у животного тяжелого эмоционального расстройства возникают либо очень сильные симпатические реакции, либо парасимпатические. В любом случае все поведение в целом становится патологическим.
Важно подчеркнуть, что внешние факторы, вызывающие значительное повышение реактивности симпатической системы, такие, как электрошок, инсулиновая кома и метразоловые судороги, ведут к сходным изменениям условных рефлексов. Несколько таких воздействий может привести к восстановлению ранее заторможенных условных рефлексов [355, 463], но серия из 10 или более приводит к устранению условной реакции страха [495] или к парадоксальным изменениям реакций на условные и дифференцировочные раздражители [764]. Исходя из этого, мы полагаем, что патологическое поведение возникает вследствие вегетативных нарушений на гипота-ламичееком уровне. Они характеризуются чрезмерно сильными симпатическими разрядами или нарушениями вегетативного баланса, возникающими под влиянием конфликтной ситуации или шоковой «терапии». Можно считаггь, что эти процессы причинно связаны и что изменения поведения в значительной степени зависят от изменения гипоталамо-кортвкальных разрядов.
ТЕОРИИ ЭМОЦИЙ
ПРОЦЕССЫ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Наше стремление подчеркнуть важное значение гипоталамуса и висцерального мозга для эмоций не исключает рассмотрения активации нервного аппарата в более широком аспекте.
Важные открытия последних лет показывают, что некоторые части центральной нервной системы оказывают диффузное возбуждающее или тормозное влияние на спинной и головной мозг. Восходящая активирующая ретикулярная система, описанная Мэтуном и другими [582, 584, 860, 861], которая при раздражении вызывает бодрствование и десинхронизацию в ЭЭГ, может служить классическим примером возбуждающей системы. Сходные эффекты можно получить с заднего отдела гипоталамуса ,и различных частей висцерального мозга. Рефлекторное возбуждение гипоталамуса через болевые и проириоцептивиые нервы [59] или высвобождение заднего отдела гипоталамуса из-под ограничивающих влияний барорецепторов синуса аорты аналогичным образом вызывает реакцию пробуждения в ее поведенческом и электроэнцефалографическом выражении [674]. Диффузная активация .коры, сопровождаемая реакцией пробуждения, возникает также при раздражении медиальных ядер таламуса и хвостатого ядра, если частота раздражения достаточно высока (100 имп/сек). Поскольку эти структуры усиливают также движения, вызываемые раздражением моторной коры, и повышают спинномозговые рефлексы [702], можно сказать, что в мозге существует возбуждающая система, к которой относятся корковые и подкорковые структуры и которая активирует сенсорные и моторные системы. Это в равной мере относится к реакции пробуждения и к эмоциональным реакциям: они могут быть вызваны с корковых и подкорковых структур и усиливают движения [671] и сенсорные процессы (восприятие [307]).
Напротив, существует тормозная система, снижающая двигательную активность и уровень активности коры. К ней относится ретикулярная формация продолговатого мозга, оказывающая тормозное влияние на спин-
18-517
ГЛАВА XXV
|номозго1вые рефлексы [617]. Кроме того, известно, что некоторые ядра мозга, например интраламинарные ядра ташамуса [186] и хвостатое ядро [830], оказывают синхронизирующее влияние на кору больших полушарий и подкорковые структуры, особенно при низкочастотном I раздражении или раздражении одиночными ударами. При этих условиях раздражение таламуса вызывает
(«реакцию остановки» (подобную временной потере сознания во время эпилептического припадка); раздражение хвостатого ядра снижает спонтанную активность [9] и подавляет движения, вызванные раздражением коры [640]. Раздражение гиппокамиа током низкого напряжения [507] и переднего отдела гипоталамуса [915] также может вызвать синхронизацию в коре. Последние работы показали еще более широкое распространение тормозной системы в мозге. Исследования ЭЭГ животных после перерезки ствола мозга кпереди от места вхождения V пары черепномозговых нервов (претригеми-нальный оредиемостовой препарат) показали, что синхронизирующие влияния могут возникать в каудальной части ствола мозга [156, 624], возможно в тормозном центре продолговатого мозга, по терминологии Мэгуна и Райнеса [617].
Значение этой работы очевидно из того факта, что сон можно вызнать не только раздражением гипниген-ной зоны Гесса, тесно связанной с интраламинарными ядрами таламуса, но также и переднего отдела гипоталамуса [34] и даже ретикулярной формации среднего мозга [139]. Интересно, что раздражение одного и того же участка в ретикулярной формации вызывает у обезьян зевоту и сон при напряжении 0,2 в, пробуждение при 0,4 в и выраженную паническую реакцию с признаками максимального возбуждения симпатической системы при 0,65—0,9 в [727]. Характер раздражения часто более важен, чем место его нанесения. Так, низкочастотное раздражение хвостатого ядра и интраламииарных ядер таламуса вызывает в коре синхронизацию, а высо-. кочастотпое — десинхронизацию.
Интересно, что процессы возбуждения сопровождаются признаками активации симпатической системы, тогда как тормозные состояния связаны с признаками
ТЕОРИИ ЭМОЦИИ
повышенной парасимпатической активности [394, '„926]. Эти данные показывают, что в функциональном отношении (Представления об эрго- и трофотропной системах могут быть расширены за пределы гипоталамуса. Несомненно, смена сна и бодрствования, а также различные эмоции сопровождаются сдвигом равновесия между возбуждающей и тормозной системами, поскольку между передним >и задним отделами гипоталамуса существуют реципрокные отношения, причем приятные эмоции в основном, очевидно, связаны с передним отделом гипоталамуса, а неприятные главным образом с задним. Яркой иллюстрацией этого может служить тот факт, что у кошки поглаживание по спине, вызывающее приятные ощущения, сопровождается синхронизацией в коре (подобно тому что имеет место при раздражении переднего отдела гипоталамуса), повышением возбудимости переднего отдела гипоталамуса и торможением тонических разрядов гамма-эфферентной системы [916]. Эти три явления объяснимы как результат парасимпатической настройки гипоталамуса и сопутствующего реципрокно-го торможения его симпатического отдела.
Поскольку эмоциональные расстройства часто приводят к грубой патологии, возникает вопрос, не ограничивает ли тормозная система: активность возбуждающей системы, и наоборот? Чтобы ответить на этот вопрос, надо проанализировать реактивность тормозной системы при разных уровнях активности возбуждающей системы.
Дата добавления: 2014-12-10; просмотров: 1540;