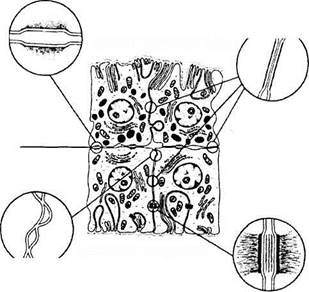
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА. Рис. 3.8.29.Топографические особенности ресничного тела:


Рис. 3.8.29.Топографические особенности ресничного тела:
/ — зубчатая линия; 2 — ресничный кружок; 3— ресничный венец; 4 — корень радужной оболочки; 5 — хрусталик
снизу и темпорально (5,6—6,3 мм), а уже — вверху и назально (4,6—5,2 мм) [959]. Необходимо помнить, что размер ресничного тела прямо коррелирует с передне-задним размером глаза. Так, при буфтальме оно достигает 10 и более миллиметров.
Граница между ресничным телом и сетчаткой при транссклеральной диафаноскопии выглядит темной полосой, поскольку с обеих сторон зубчатой линии пигментный эпителий интенсивно окрашен. Ширина этой полосы — 1,5—2,0 мм. Передняя часть полосы шириной 1,0—1,5 мм принадлежит ресничному телу, что необходимо учитывать при хирургическом проникновении в глазное яблоко через ресничное тело.
Ресничное тело легко механически отделить от склеры. Для этого достаточно пересечь место его прикрепления к склеральной шпоре. Поверхность склеры, соответствующая локализации ресничного тела, гладкая и слабо пигментирована. Видны лишь каналы, через которые проникают ресничные артерии и нервы.
Ресничное тело четко разделяется на 2 части: часть, имеющую многочисленные гребешки (ресничный венец; corona ciliaris), и широкую плоскую заднюю часть (ресничный кружок; orbiculus ciliaris; pars plana). Ширина ресничного венца равняется 2,0 мм, а плоской части — 4,0—4,5 мм.
Ресничный венец состоит приблизительно из 70—80 небольших гребешков, ориентированных радиально (рис. 3.8.30; 3.8.31, см. цв. вкл.; 3.8.32). В пространстве между гребешками лежат маленькие, неравномерно пигментированные складки (ресничные складки; plicae ciliaris).
Ресничные отростки располагаются симметрично и разнообразны в размере (длина 2,0 мм; ширина 0,5 мм) [154, 496]. Иногда (16% случаев) видны гигантские отростки, чаще располагающиеся с назальной стороны [1026, 1049]. Гигантские отростки нередко сочетаются с аномалиями развития цинновой связки и наличием периферической дегенерации сетчатки.
Плоская часть ресничного тела простирается от заднего края ресничных гребешков
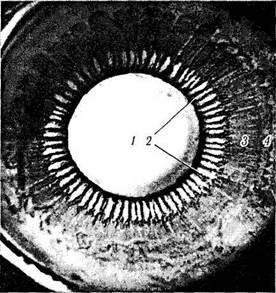
Рис. 3.8.30. Строение ресничного тела при рассмотрении его со стороны стекловидного тела:
/ — задняя поверхность хрусталика; 2— ресничный венец; 3— ресничный кружок; 4 — зубчатая линия
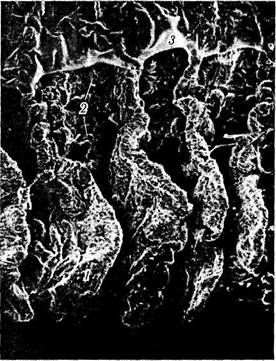
Рис. 3.8.32. Сканограмма отростков ресничного тела:
/ — большие отростки; 2 — маленькие отростки; 3 — остатки стекловидного тела, прилежащие к ресничному кружку
до зубчатой линии (4 мм). Таким образом, отношение ширины плоской и отростчатой частей ресничного тела на меридиональных срезах равно 2:1. Плоская часть ресничного тела неравномерно пигментирована. Более пигментирована она с темпоральной стороны.
Сосуды и сосудистая оболочка глазного яблока

 Плоская часть ресничного тела содержит относительно небольшое количество сосудов. По этой причине оперативное вмешательство в этой области не сопровождается массивным кровотечением и повреждением сетчатки. У новорожденных ресничное тело короче, а его плоская часть смещена кпереди.
Плоская часть ресничного тела содержит относительно небольшое количество сосудов. По этой причине оперативное вмешательство в этой области не сопровождается массивным кровотечением и повреждением сетчатки. У новорожденных ресничное тело короче, а его плоская часть смещена кпереди.
Более подробно размеры ресничного тела и его структурных образований приведены в табл. 3.8.1.
Таблица 3.8.1. Размеры ресничного тела
Длина плоской части, мм.......................................... 3,5—4,0
Длина отростчатой части, мм.............................................. 2,0
Количество ресничных отростков.............................. 70—80
Длина ресничных отростков, мм........................................... 2
Толщина ресничных отростков, мм................................... 0,5
Расстояние между склеральной шпорой и зубчатой линией, мм
темпорально........................................................ 7,5—8,0
назально............................................................... 6,5—7,0
Высота клеток непигментированного эпителия, мкм
отростчатая часть................................................... 10—15
плоская часть.......................................................... 20—30
Высота пигментированного эпителия, мкм................. 8—15
В соответствии с особенностями микроскопического строения ресничное тело можно разделить на 6 слоев (рис. 3.8.33, 3.8.34):
1)супрацилиарный слой (супрахориоидаль-
ное пространство);
2) ресничная мышца;
3) слой сосудов;
4) наружная базальная мембрана;
5) эпителий;
6) внутренняя базальная мембрана.
Основываясь на данных эмбриологии, т. е.
учитывая особенности происхождения той или иной структуры, ресничное тело может быть разделено на две части (слоя): внутренняя часть (нейроэпителиальная) и наружная (уве-альная, мезодермальная). Эту классификацию мы и будем использовать при последующем изложении материала.
Нейроэпителиальная часть(рис. 3.8.31, 3.8.33—3.8.36). Нейроэпителиальная часть ресничного тела формируется в эмбриональном периоде и представляет собой два слоя эпителиальных клеток зрительного бокала. При этом клетки эпителиальных слоев прилежат друг к другу своими апикальными поверхностями. Наружный (пигментированный) слой ресничного эпителия является продолжением пигментного эпителия сетчатки. При этом клетки этого слоя практически не отличаются от клеток пигментного эпителия сетчатки.
Клетки внутреннего слоя ресничного эпителия по происхождению аналогичны клеткам сенсорной части сетчатки. Сенсорная часть сетчатки внезапно прерывается у зубчатой линии, продолжаясь кпереди на ресничное тело, формируя внутренний беспигментный слой ресничного эпителия.
В области ресничного тела просвет зрительного пузырька полностью облитерируется. При этом два слоя эпителиальных клеток плотно сращены благодаря наличию многочисленных межклеточных контактов [154, 317](рис. 3.8.34). Попытка их механического разделения приводит к разрушению апикальной части клеток.
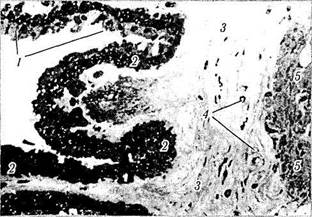
Рис. 3.8.33. Микрофотография ресничного венца ресничного тела (по Hogan et al., 1971):
I — беспигментный эпителий (внутренний слой), содержащий незначительное количество пигментных гранул; 2 — клетки пигментированного слоя (наружного), содержащие большое количество зерен меланина в апикальной части; 3— строма ресничного тела; 4 — кровеносные сосуды стромы; 5 — участок ресничной мышцы
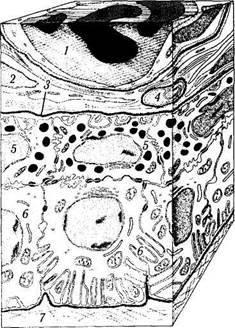
Рис. 3.8.34. Схематическое изображение строения
пигментного эпителия ресничного тела (по Poliquen,
1969):
1 — капиллярный сосуд; 2 — фибробласт; 3 — базальная мембрана; 4 — нервный ствол; 5 — наружный слой клеток пигментного эпителия; 6— внутренний слой клеток пигментного эпителия, 7 — базальная мембрана
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

|
| Точка сращения |
| Щелевой контакт Базальная поверхность пигментированного слоя |
| Апикальная поверхность |
| Базальная поверхность беспигментного слоя Плотный контакт |
Десмосома
Рис. 3.8.35. Типы межклеточных контактов, обнаруживаемых между клетками пигментного эпителия ресничного тела (по Raviola et al., 1977)

Рис. 3.8.36. Электроннограмма эпителия ресничного тела:
/—внутренняя пограничная мембрана; 2—митохондрии; 3— гладкий эндоплазматический ретикулум; 4 — аппарат Гольджи; 5 — цитоплазматические отростки, распространяющиеся в подлежащую строму пигментных эпителиальных клеток; 6 — базальная мембрана пигментных эпителиальных клеток; 7 — меланино-вые гранулы пигментированного слоя эпителия
Кроме указанных контактов между двумя эпителиальными клеточными слоями обнаруживается и третий компонент, участвующий в межклеточном соединении — межклеточная цементирующая субстанция [317]. Она расположена внеклеточно между апикальными поверхностями клеточных мембран и имеет вид сплошной линии.
Беспигментный слой пигментного эпителия и внутренняя базальная мембрана. Внут-ренная базальная мембрана ресничного тела располагается с базальной (витреальной) стороны беспигментных клеток эпителия. Она связана с внутренней пограничной мембраной сетчатки сзади и радужки — спереди.
У новорожденных внутренняя базальная мембрана имеет типичное строение базальной мембраны. Состоит она из зернистого слоя толщиной 30 нм и прозрачного слоя (lamina lucida), толщина которого равна 50 нм. Состоит она из коллагеновых волокон I, III и IV типов, а также ламинина.
В возрасте 3 лет базальная мембрана начинает утолщаться и становится многослойной.
По направлению базальной мембраны ба-зальные поверхности беспигментных клеток образуют многочисленные складки и отростки, которые оплетаются волокнами мембраны и коллагеновыми волокнами основания стекловидного тела.
Беспигментные эпителиоциты имеют кубическую форму в отростчатой части ресничного тела (ширина 12—15 мкм и высота 10—15). В плоской части ресничного тела они цилиндрические (ширина 6—9 мкм и высота 30 мкм). Наиболее высокие клетки лежат вблизи зубчатой линии, возможно, в результате их тракции стекловидным телом. Многими исследователями отмечено увеличение высоты клеток с возрастом.
Между лежащими по соседству беспигментными эпителиальными клетками обнаруживаются щелевидные пространства различной ширины [317]. Эти пространства заполнены прозрачным внеклеточным муцинозным материалом, положительно окрашивающимся при проведении гистохимических реакций, направленных на выявление кислых гликозаминогли-канов.
Цитоплазма эпителиоцитов насыщена органоидами, что указывает на их высокую секреторную функцию. Особенно много в цитоплазме митохондрий [443]. Митохондрии являются основным источником энергии для обеспечения активного транспорта электролитов и молекул небольшого размера через цитоплазму эпителиальных клеток в процессе формирования камерной влаги.
В эпителиальных беспигментных клетках также хорошо развит шероховатый и гладкий эндоплазматический ретикулум [317, 496]. Цитоплазма эпителиоцитов содержит большое ко-
Сосуды и сосудистая оболочка глазного яблока
 личество промежуточных микрофиламентов и филаментов актина. Необходимо отметить, что микрофиламенты положительно окрашиваются при выявлении виментина и цитокератана.
личество промежуточных микрофиламентов и филаментов актина. Необходимо отметить, что микрофиламенты положительно окрашиваются при выявлении виментина и цитокератана.
Боковые поверхности беспигментных клеток образуют многочисленные складки цитоплазма-тической мембраны, погруженные в цитоплазму соседних клеток (интердигитации). Складки значительно увеличивают площадь контакта между клетками, что наиболее выражено в передней части ресничных отростков [154, 795, 872, 875, 890, 1125].
Апикальные поверхности беспигментных эпи-телиоцитов ресничных отростков при макроскопическом исследовании белые, что связано с высоким содержанием в их цитоплазме гликогена.
На переднем склоне ресничных гребешков по мере приближения к радужной оболочке в части эпителиальных клеток появляются зерна меланина. Степень пигментации постепенно нарастает, пока они не переходят в слой пигментного эпителия корня радужки.
Хотя внутренний слой ресничного эпителия и называется непигментированным, с возрастом возможно появление в их цитоплазме зерен меланина и липофусцина.
Большое значение в функционировании эпителия ресничного тела имеют межклеточные контакты, расположенные как между беспигментными клетками, так и между беспигментными и пигментированными клетками [872, 874, 875] (рис. 3.8.34, 3.8.35). Наиболее важным в функциональном смысле типом контакта является так называемая лента замыкания, располагающаяся на боковых поверхностях беспигментных клеток в апикальной их части. Встречаются также контакты типа «кепки», соединяющие боковые поверхности пигментированных и реже беспигментных клеток, щелевые контакты и десмосомы. Особое внимание в настоящее время уделяется изучению щелевых контактов. Предполагают, что эти контакты обеспечивают существование так называемого «электрического» взаимодействия эпителиальных клеток между собой. Именно подобный тип взаимодействия позволяет эпителиальному пласту функционировать как синцитий и, возможно, обеспечивает координацию секреторной деятельности всего эпителиального пласта.
Наличие перечисленных контактов между клетками, особенно ленты замыкания, делает практически невозможным проникновение между клетками высокомолекулярных веществ путем диффузии [1016].
Как неоднократно указывалось выше, беспигментный слой эпителиальных клеток ресничного тела является основным структурным элементом, обеспечивающим секрецию камерной влаги. Вышеприведенные светооптические, ультраструктурные особенности клеток, набор органоидов и плотность межклеточных контактов способствуют этому процессу. Из кровяно-
го русла капилляров ресничных отростков в переднюю камеру ионы и вещества небольшой молекулярной массы проходят благодаря функционированию энергозависимой транспортной системы [205, 206]. В соответствии с Cole [205] непигментированные клетки ресничного эпителия выборочно поглощают ионы натрия из стромы ресничного тела и транспортируют их через межклеточные пространства. Гиперосмо-тичность, возникающая в межклеточных пространствах в результате этого процесса, приводит к осмотическому потоку воды первоначально из стромы в межклеточные пространства, а затем в камерную влагу. О правильности подобного предположения свидетельствует обнаружение в мембранах непигментированных клеток ресничного эпителия ряда ферментов, участвующих в этом процессе, в частности активности Na+/K+-ATO-a3bi, угольной ангидразы [154, 327, 674].
Подтверждением тому являются и данные, относительно того, что применение средств, ин-гибирующих активность №+/К+-АТФ-азы, уменьшает секрецию камерной влаги. Прохождение ионов хлорида, бикарбоната и калия также обеспечивается активным транспортом. Ряд веществ проходит через эпителий пассивно, благодаря перепаду градиента концентрации. Это относится к сахарам и аминокислотам [241].
Секреция бикарбонатов снижается при применении ингибиторов угольной ангидразы (кар-боангидраза), что и используют при глаукоме для снижения секреции камерной влаги. Такими же свойствами обладают бета-блокаторы.
Беспигментные эпителиоциты ресничного тела, помимо секреции камерной влаги, участвуют еще во многих процессах.
Они синтезируют компоненты стекловидного тела — коллагены различного типа и глюку-роновую кислоту [124, 133, 716]. Участвуют эти клетки и в синтезе неколлагеновых белков — оптицина, фибулина-1 и нидогена-1, играющих немаловажную роль в стабилизации геля стекловидного тела и организации базальных мембран [717, 879].
Помимо основных секретирующих функций беспигментный эпителий принимает участие во многих репаративных и адаптационных процессах. Так, совсем недавно установлено, что он способен ингибировать пролиферативную активность и функции Т-лимфоцитов [1213], регулируя тем самым иммунные механизмы внутри глазного яблока.
Пигментированный эпителий и наружная базальная мембрана (рис. 3.8.32—3.8.36). Как было указано выше, пигментный слой эпителия ресничного тела развивается из наружного слоя зрительного бокала. По направлению к задним отделам глаза он переходит в пигментный эпителий сетчатой оболочки, а по направлению кпереди в передний слой пигментного эпителия радужки.
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

 Размер пигментированных эпителиальных клеток колеблется от 8—10 мкм в плоской части ресничного тела, до 10—15 мкм в ресничных отростках [959].
Размер пигментированных эпителиальных клеток колеблется от 8—10 мкм в плоской части ресничного тела, до 10—15 мкм в ресничных отростках [959].
В цитоплазме клеток обнаруживаются крупные круглые зерна меланина (0,8—2,0 мкм, т. е. в 3—4 раза больше, чем те в клетках сосудистой оболочки). Видны также пучки микро-филаментов промежуточного типа [285]. Цитоплазма клеток насыщена органоидами [747].Базальная мембрана пигментных эпителиоци-тов образует многочисленные складки. Таким образом, микроскопически эти клетки практически не отличаются от клеток пигментного эпителия сетчатки.
Увеальная (мезодермальная) часть.Уве-альная часть ресничного тела имеет сложное строение и состоит из сосудов, мышечной ткани и стромы. К увеальной части относится также наружная базальная мембрана. На перечисленных структурных элементах мы сейчас остановимся более подробно.
Слой сосудов увеальной части ресничного тела является продолжением сосудистого слоя хориоидеи. Между ним и базальнои мембраной эпителия лежит слой коллагеновых волокон большого диаметра. По мере продвижения кпереди сосудистая ткань как бы собирается в складки, формируя соединительнотканную основу ресничных отростков.
Строма ресничного тела состоит из волокон, клеток и межклеточного вещества. Основными типами клеток стромы являются фибро-бласт и меланоцит. Обнаруживаются также моноциты, макрофаги, лимфоциты и дендритические клетки.
В последние годы обращено особое внимание на изучение функции моноцитов, макрофагов и дендритических клеток. Это связано с тем, что названные клетки определяют иммунную резистентность тканей глаза. Именно макрофаги сталкиваются с чужеродным антигеном, распознают его и передают информацию другим элементам иммунной системы.
Как и макрофаги других органов и тканей, макрофаги ресничного тела обладают необходимым набором рецепторов опознания чужеродного антигена, что показано при использовании методов иммуноморфологии [581, 724—726]. Более того, при воспроизведении экспериментального увеита или возникновении заболевания у человека в моноцитах и макрофагах ресничного тела и сосудистой оболочки происходит вся цепь структурных и функциональных изменений, свойственных макрофагам других локализаций при встрече с чужеродным антигеном [168].
Основной объем ресничного тела занимает соединительнотканная строма. В зависимости от расположения различают несколько слоев.
Внутренний слой соединительной ткани находится между мышцей и базальнои мембраной
пигментного эпителия. Этот слой довольно тонкий в плоской части ресничного тела, но утолщается в отростчатой части. В поверхностных слоях коллагеновые волокна этого слоя смешиваются с волоканами более плотной соединительной ткани ресничной мышцы. По направлению кпереди этот волокнистый слой переходит в строму радужной оболочки.
Передний слой соединительной ткани ресничного тела располагается на границе между ресничной мышцей и передней камерой глаза (рис. 3.8.39).
Строма ресничного тела несколько отличается от плотной неоформленной соединительной ткани, обнаруживаемой в других местах организма человека. Эти отличия сводятся, в первую очередь, к структуре эластических волокон. Эластические волокна на поперечном разрезе имеют вид трубочек, напоминая при этом волокна зонулярного аппарата [1048, 1053]. При этом эластические волокна формируют довольно густую сеть, в которую и помещены мышечные волокна. Иммуногистохимически выявляется большое количество микрофибриллярного белка фибриллина [1162].
Определенные отличия выявлены и в отношении преобладающего типа коллагена, из которого состоят коллагеновые волокна стромы. Наиболее распространен коллаген VI типа, хотя выявляются и коллагены других типов [895]. Коллаген VI типа преимущественно локализуется вокруг капиллярных сосудов. При этом количество данного типа коллагена больше со стороны, обращенной к эпителию. Коллаген VI типа образует также оболочки передних сухожилий ресничной мышцы, направляющихся к трабекулярной сети [671, 675, 909].
Коллагеновые и эластические волокна окутывают мышечные волокна, образуя межмышечную соединительную ткань. Именно указанное сочетание мышечных, коллагеновых и эластических волокон образует жесткую систему, передающую силу, возникающую при сокращении мышечных клеток, так называемым «сухожилиям», распространяющимся в ресничном теле особым образом (см. ниже).
В заключение необходимо указать на значительное количество в строме ресничного тела нервных стволов и кровеносных сосудов различного калибра (рис. 3.8.33, 3.8.37, 3.8.38).
Мембрана Бруха ресничного тела.Кути-кулярный слой мембраны Бруха продолжается на ресничное тело в виде базальнои мембраны пигментного эпителия. Эластическая и колла-геновая части мембраны Бруха в области сосудистой оболочки отделяются от базальнои мембраны слоем коллагена и, в конечном счете, исчезают в строме ресничного тела вблизи плоской части.
Ресничная мышца(т. ciliaris) (рис. 3.8.39— 3.8.42). Особое место, как в структурном, так и функциональном отношениях, занимает рее-
Сосуды и сосудистая оболочка глазного яблока

|
а 6
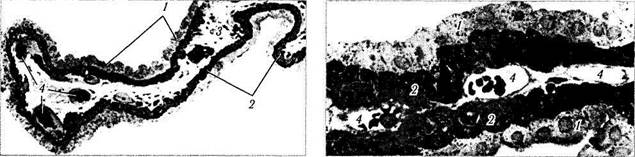
Рис. 3.8.37. Ресничный отросток (а — малое увеличение; б — большое увеличение):
/ — непигментированный ресничный эпителий; 2—пигментированный ресничный эпителий; 3— строма; 4— капиллярные сосуды,
часть которых прилежит непосредственно к ресничному эпителию
|
| шшшш |
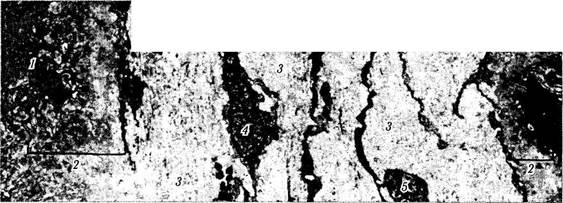
Рис. 3.8.38. Строма ресничного отростка ресничного кружка (по Hogan, I972):
базальная часть пигментного эпителия видна слева (/) и прилежит к толстой базальной мембране (2). Стромальный коллаген (3) состоит из плотных пучков коллагеновых волокон, ориентированных в различных направлениях. Видны также фибробласты (4)
и нервные волокна (5)
ничная мышца. Именно благодаря деятельности мышцы осуществляется аккомодация глазного яблока.
Гладкомышечные волокна ресничного тела располагаются таким образом, что при своем сокращении в трех направлениях происходит деформация ресничного тела. Соответственно различают три группы волокон:
1. Наружные меридианальные (продольные)
пучки волокон (fibrae meridionales) (мышца
Брюкке), составляют большую часть реснич
ной мышцы. Начинаются они в подсосудистой
пластинке вблизи ресничного тела и проходят
до склеральной шпоры, прикрепляясь над тра-
бекулярной сеточкой.
2. Радиальные пучки волокон (fibrae radla-
les), лежащие кнутри меридианальных. На
правляются они изнутри кнаружи, пересекая
под прямым углом меридианальные и циркуляр
ные волокна ресничной мышцы. Распростра
няясь назад, они прикрепляются на широком
участке к соединительной ткани сосудистой
оболочки.
3. Циркулярные пучки волокон (fibrae
radiales) (мышца Мюллера). Лежат такие во-
локна у внутреннего края ресничного тела вблизи его основания, кнутри от меридианальных волокон. Волокна циркулярно окружают ресничное тело.
Основная масса ресничной мышцы располагается в передних 2/3 ресничного тела. Сокращение среднего и наружного слоев ресничной мышцы в наибольшей степени приводит к смещению ресничных отростков кпереди и кнутри [772] и расслаблению ресничного пояска.
Мышечные волокна ресничного тела, хотя и относятся к гладкомышечным волокнам, тем не менее обладают и определенными особенностями строения. Их цитоплазма содержит большое количество митохондрий. Исключительно хорошо развита эндоплазматическая сеть. Аппарат Гольджи находится в активном функциональном состоянии.
Определенные отличия от гладкомышечной ткани других органов существуют и в характере связи между отдельными мышечными клетками. Мышечные клетки ресничного тела складываются в так называемые «связки», окруженные тонким слоем фибробластов (рис. 3.8.39, 3.8.40). В определенных точках мышечные клет-
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА


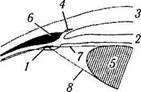
Рис. 3.8.39. Меридианальный срез ресничного тела:
/ — продольный пучок ресничной мышцы, прикрепляющейся к склеральной шпоре; 2— склеральная шпора; 3— циркулярный пучок ресничной мышцы; 4 — угол передней камеры; 5 — сосуд большого круга кровообращения радужной оболочки; 6 — радиальный пучок ресничной мышцы; 7 — пигментный эпителий ресничного тела

Рис. 3.8.40. Электроннограмма продольного среза ресничной мышцы (по Hogan et al., 1971):
I — базальная мембрана, окружающая мышечные клетки; 2 —
десмосомоподобные уплотнения, расположенные на внутренней
поверхности цитоплазматической мембраны мышечных клеток;
3 — миофиламенты
ки прикрепляются друг к другу при помощи десмосом. Группы мышечных клеток окружены тонким слоем коллагеновой ткани и перимизием (рис. 3.8.40).
Как и все гладкомышечные клетки организма человека, миоциты ресничного тела содержат миофиламенты, преимущественно располо-
женные по периферии клетки. При этом основной особенностью миофиламентов является обнаружение в них структурных признаков поперечнополосатой мышцы, поскольку они располагаются практически параллельно друг другу и прикрепляются к электронноплотным структурам цитоплазматической мембраны, напоминающим Z-связки.
Необходимо отметить и то, что, как и поперечнополосатая мышца, ресничная мышца исключительно хорошо иннервирована [1085].
В этом разделе уместно остановиться и на основной функции ресничной мышцы, а именно на ее участии в процессе аккомодации.
Аккомодация является процессом фокусировки изображения на сетчатую оболочку, а дисаккомодация представляет собой обратный процесс (расфокусировка). Основную роль при этом играет способность хрусталика к обратимой деформации в процессе сокращения и расслабления ресничной мышцы и сопровождающие этот процесс расслабление и натяжение цинновой связки хрусталика. Именно поэтому происходит деформация хрусталика.
Аккомодация сопровождается сужением зрачка, смещением радужки кпереди, увеличением кривизны передней и, в меньшей степени, задней поверхностей хрусталика, увеличением толщины хрусталика [157, 158, 162, 607, 964—966].
Вопросы функции ресничной мышцы в процессе аккомодации наиболее полно изучались у обезьян [904]. Существуют доказательства того, что выявленные у обезьян механизмы аккомодации распространяются и на человека. Первоначально необходимо остановиться на особенностях взаимоотношения ресничной мышцы с окружающими структурами и, в частности, с хрусталиком.
Целесообразно начать с данных, относительно особенностей прикрепления «сухожилий» ресничных мышц. При этом различают передние и задние «сухожилия».
Передние «сухожилия», начинаясь от мышц, расходятся в виде веера, разделяясь на три части. Одна часть сухожильных волокон прикрепляется к передней части склеры, вторая — к склеральной шпоре, а третья — вплетается в волокнистую часть трабекулярного аппарата.
Задние «сухожилия» ресничной мышцы прикрепляются в другом месте, а именно в области плоской части ресничного тела. В отличие от передних «сухожилий» задние «сухожилия» содержат большое количество эластической ткани и вплетаются в адвентицию кровеносных сосудов, эластический слой мембраны Бруха ресничного тела, а также базальную мембрану ресничного эпителия.
Вышеприведенные особенности прикрепления передних и задних «сухожилий» создают систему, которая при сокращении или расслаблении ресничной мышцы быстро приводит к де-
Сосуды и сосудистая оболочка глазного яблока

|
| формации и смещению ресничного тела в раз- ки хрусталика в плоскости экватора [300, 901, |
| 904, 911] (рис. 3.8.42). С этим и связывают различия в степени изменения кривизны передней и задней поверхностей хрусталика. |
личных направлениях [154, 162, 901, 904, 608, 964-966, 1052, 1067].

|
| Зубчатая линия |
При синхронном сокращении всех частей мышцы диаметр «кольца» ресничного тела уменьшается. При этом внутренняя граница мышцы перемещается к хрусталику. Уменьшение диаметра «мышечного кольца» приводит к расслаблению цинновой связки, снижая напряжение капсулы хрусталика. В результате своей упругости, хрусталик изменяет форму, становясь более выпуклым [314] (рис. 3.8.41). Передний полюс хрусталика при этом перемещается вперед, а задний остается на месте или слегка перемещается назад. Эти изменения конфигурации хрусталика и его перемещение и увеличивают силу рефракции.
Рис. 3.8.41. Деформация ресничного тела и подтягивание кпереди зубчатой линии при сокращении ресничной мышцы:
а — сокращение ресничной мышцы; б — расслабление ресничной мышцы
Необходимо отметить, что в процессе аккомодации не изменяется кортикальная толщина хрусталика, но увеличивается сагиттальная толщина ядра. Именно благодаря этому происходит утолщение хрусталика в целом.
Много исследований было посвящено выяснению причин различной степени изменения кривизны передней и задней поверхностей хрусталика. Известно, что места прикрепления передних и задних зонулярных волокон цинновой связки к капсуле хрусталика различны (см. «Хрусталик»), Поэтому сила напряжения зонулярных волокон направлена радиально от ресничного тела в направлении разветвления связ-

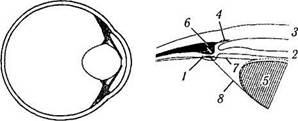
В
Рис. 3.8.42. Схема изменения геометрии глаза при расслаблении ресничной мышцы (о, б) и аккомодации (в, г) (по Rohen, 1979):
I — система фибрилл зонулярного аппарата; 2 — радужная оболочка; 3 — роговица; 4 — шлеммов канал; 5 — хрусталик; 6 — ресничная мышца; 7 — волокна передней части цинновой связки; 8 — волокна задней части цинновой связки. Стрелка указывает направление движения ресничной мышцы в процессе аккомодации. Ресничная мышца при сокращении смещает внутренний край ресничного тела по направлению экватора хрусталика. При этом волокна передней цинновой связки расслабляются и хрусталик принимает более сферическую форму (пунктирная линия). Сосудистая оболочка подтягивается к центру и кпереди
Другие исследователи считают, что различная степень изменения кривизны передней и задней поверхностей хрусталика связана с неодинаковой толщиной капсулы хрусталика в различных местах [314].
Coleman [207, 208] предложил «гидравлическую теорию». По его мнению, меньшее изменение кривизны задней поверхности хрусталика при аккомодации связано с наличием давления на него стекловидного тела. Тем не менее Fisher [323] предполагает, что стекловидное тело не влияет на этот процесс. По его мнению, способность хрусталика к деформации всецело зависит от его физических свойств. Причем способность к деформации существенно отличается в центральной и экваториальной плоскостях хрусталика.
При сокращении ресничной мышцы происходит и ряд других структурных изменений в переднем отделе глаза. Так, сокращение части мышцы, прикрепляющейся к склеральной шпоре, приводит к расширению межтрабекулярных пространств, что способствует усилению фильтрации камерной влаги [412, 413, 676—679].
Дата добавления: 2014-12-20; просмотров: 1059;