Морфология и тинкториальные свойства
МБТ — грамположительные прямые или слегка изогнутые неподвижные палочки, патогенные для человека и многих видов животных. Наиболее восприимчивы к заражению морские свинки.
МБТ по форме напоминают палочки длиной 1 — 10 мкм (чаще 1-4 мкм), шириной 0,2-0,7 мкм; по виду они могут быть гомогенными или зернистыми со слегка загнутыми краями.
Электронно-микроскопические исследования МБТ позволили дифференцировать в них микрокапсулу, многослойную клеточную мембрану, цитоплазму с органеллами (гранулы, вакуоли, рибосомы) и ядерную субстанцию (нуклеотид) (рис. 2.1).
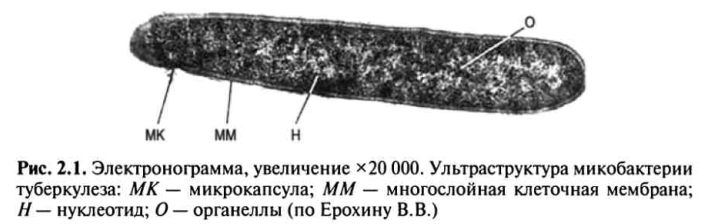
Микрокапсула состоит из полисахаридов и играет важную роль в жизнедеятельности МБТ, в том числе придает им устойчивость к неблагоприятным воздействиям внешней среды.
Толстая клеточная стенка, насыщенная липидами, ограничивает клетку снаружи, обеспечивая механическую и осмотическую защиту (рис. 2.2).

Клеточная стенка МБТ состоит из четырех слоев. Первый (внутренний) слой образован пептидогликаном; последующие слои включают миколевые кислоты, гликолипиды, воск и корд-фактор (фактор вирулентности), которые обусловливают склеивание МБТ и оказывают токсическое действие на макрофаги при фагоцитозе. При этом высоковирулентные штаммы МБТ на твердых питательных средах образуют культуры в виде кос и жгутов (рис. 2.3, см. цветную вклейку).
Такой состав клеточной стенки определяет устойчивость МБТ к воздействию кислот и щелочей, а также высокую гидрофобность клетки в целом.
Согласно современным представлениям, в состав цитоплазматической мембраны, расположенной под клеточной стенкой, входят липопротеидные комплексы. С мембраной связаны ферментные системы. В цитоплазматической мембране осуществляются процессы, ответственные за специфичность реакций микобактериальной клетки на окружающую среду.
Цитоплазма МВТ состоит из гранул и вакуолей различной величины. Основная часть мелкогранулярных включений представлена рибосомами, на которых синтезируется специфический белок.
Ядерная субстанция МВТ определяет специфические свойства клетки, причем важнейшие из них — синтез белка и передача наследственных признаков потомству.
Для нормального развития МВТ нуждаются в кислороде, поэтому их относят к аэробам. Потребление кислорода микробной клеткой связано с окислительно-восстановительными процессами и высоким уровнем синтеза аденозинтрифосфорной кислоты (АТФ) в МВТ. При формировании гранулем размножение МВТ замедляется из-за снижения в них парциального давления кислорода. Однако имеются сведения о том, что некоторые виды МВТ можно рассматривать как факультативные анаэробы.
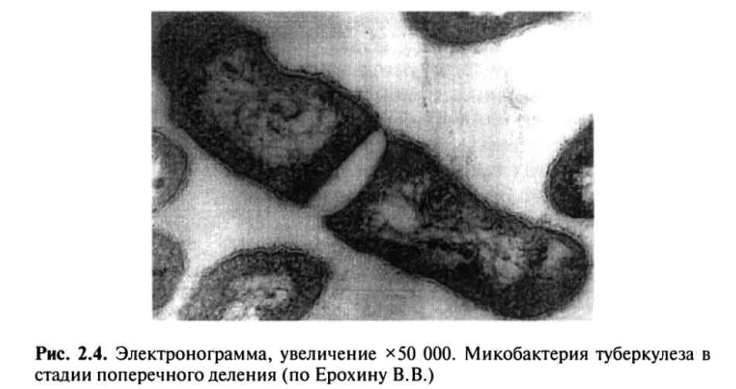
Физиологические свойства.Размножение МВТ происходит путем простого деления клетки. При электронно-микроскопическом исследовании установлено, что деление начинается с двустороннего вдавлення цитоплазматической мембраны в цитоплазму. При соединении этих перегородок образуются две дочерние особи.
Цикл простого деления материнской клетки на две дочерние занимает от 13—14 до 18—24 ч. Микроскопически видимый рост микроколоний на жидких средах можно обнаружить на 5—7-й день, видимый рост колоний на поверхности твердой среды — на 12—20-й день (рис. 2.4).
Известен более сложный цикл деления —почкование. На определенном этапе крупное гранулярное образование на одном из полюсов клетки, окруженное небольшой частью цитоплазмы, начинает перемещаться к периферии клетки, образуя выпуклость на поверхности клеточной мембраны. В дальнейшем бугорок увеличивается в размерах и отпочковывается от материнской клетки в виде образования кокковидной формы. Внутри этих форм происходит ряд превращений, в результате которых в них постепенно формируются обычные кислотоустойчивые палочки. Весь цикл размножения и воспроизводства продолжается примерно 7—9 сут.
Еще один из возможных способов размножения МВТ — спорообразование, что роднит их с актиномицетами. При этом процессе в цитоплазме клетки увеличиваются некоторые шаровидные структуры, имеющие вид зерен. Цитоплазма медленно исчезает, зерна освобождаются из клетки и могут в дальнейшем прорастать в новые особи. У МВТ выделены 2 гена (sigF и whiB) образования спор, действующих в латентной фазе роста возбудителя.
Генетическая характеристика. В 1998 г. полностью расшифрован геном МВТ. Он имеет длину 4 411 529 пар нуклеотидов, которые почти в 70% представлены гуанином и цитозином. Нуклеотид содержит 4000 генов, из них 60 кодируют компоненты рибонуклеиновой кислоты (PH К). Для МВТ имеются уникальные гены, в частности гены mtp40 и mpb70, которые применяются для выявления МВТ с помощью полимеразноцепной реакции (ПЦР) (рис. 2.5, см. цветную вклейку).
Геном МБТ радикально не отличается от других видов микобактерий. Большая часть генов обеспечивает синтез ферментов липогенезиса (синтеза миколевой кислоты, гликолипидов и воска — для построения клеточной стенки) и липолизиса (синтеза кордфактора — для разрушения клеточных структур организма человека и животных).
Тождественность дезоксирибонуклеиновой кислоты (ДНК) комплекса МБТ (Л/, tuberculosis, М. bovis, М. bovis BCG, М. africanum, М. microti, М. Canettii, М. сарrае и М. pinnipedii) составляет 99,9%.
Расшифровка геномов разных видов микобактерий позволила выделить в геноме комплекса МБТ уникальную область — RD1 (region of difference 1), отсутствующую у М. bovis и М. bovis BCG.
В дальнейшем было установлено, что данная область RD1 присутствует также у М. leprae и у ряда НТМ Б (М. kansasi и М. marinum).
В области RD1 кодируется синтез двух секреторных белков:
• ESAT-6 (early secreted antigenic target) — ранний секретируемый белок делеционного региона-6 RD1 МБТ;
• CFP-10 (culture filtrate protein-10) — белок фильтрата культуры делеционного региона-10 RD1 МБТ.
Белки ESAT-6 (с молекулярной массой около 6 кДа) и CFP-10 (с молекулярной массой 10 кДа) определяют скорость размножения МБТ, степень вирулентных свойств и массивность развития специфических изменений в органах. При этом белок ESAT-6 проявляет усиливающие свойства в комплексе ESAT-6-CFP-10, который формирует длительный постинфекционный иммунный ответ.
В то же время область RD1 отсутствует не только у М. bovis BCG и М. bovis, но также у целой группы НТМБ, таких как М. aviuv complex (MAC — М. avium, М. inracellulare, М. paratuberculosis и М. silvaticum), M. fiortuitum complex (M. fortuitum и M. chelonae), M. kansasii, M. marinum, M. scrofulaceum, M. abscessus, M. gordone, M. malmoense, M. smegmatis, M. branderi, M. celatum, M. genavense, M. terrae, M. vaccae.
При этом отсутствие области RD1, а следовательно, отсутствие возможности синтеза секреторных белков (ESAT-6 и CFP-10) М. bovis BCG позволяет проводить диагностику инфекционной и послевакцинной чувствительности к туберкул и ну у детей и подростков. Однако при отсутствии области RD1 у большого числа НТМБ, широко распространенных во внешней среде, могут возникать существенные трудности в диагностике, особенно в регионах, где распространены данные микобактерии.
На основании этих белков в Российской Федерации разработан препарат диаскинтест, который при внутрикожной пробе позволяет проводить дифференциальную диагностику поствакцинной и инфекционной аллергии у детей и подростков.
Дата добавления: 2019-10-17; просмотров: 2602;