Нервная клетка, нервное волокно, глия, синапс
Изобретение в XVII в. микроскопа позволило проникнуть в тайну строения живой и мертвой природы. Многочисленные исследования тканей, составляющих растительные и животные организмы, с помощью микроскопа показали, что они построены из мельчайших ячеек — клеток. Открытие клеточного строения живых организмов позволило выяснить некоторые сложные и неясные вопросы биологии и медицины. Боль-
Учение о клетке в дальнейшем развивалось в острых противоречиях. Спорным оказался ряд положений немецкого ученого Р- Вирхова, в течение ряда лет владевшего умами своих современников-врачей. Р. Вирхов, касаясь вопросов о путях клеткообразования, утверждал, что клетки образуются только из клеток, путем их деления. Другие пути клеткообразования отрицались. Это положение не разъясняло, а запутывало известный вопрос, являющийся предметом научного спора между материалистами и идеалистами о причинах возникновения жизни на Земле. Сущность этого спора в основном сводилась к следующему. Если живые клетки могут развиваться только из им подобных, то, естественно, возникал вопрос: как же возникла первая живая клетка, послужившая началом развития живого на Земле?
Наш мозг состоит из огромнейшего количества клеток. В одной коре больших полушарий насчитывают до 14 млрд. нервных клеток. Нервные клетки были открыты независимо от нервных волокон. Связь между нервными клетками и нервными волокнами предполагали многие исследователи, но ввиду несовершенства техники не могли ее доказать. Первые гистологические доказательства того, что нервное волокно представляет собой отросток нервной клетки, лежащей в центральной нервной системе, приводятся в работах русских ученых Ф.М. Овсянникова и Н.М. Якубовича. Позднее другим исследователям, применившим метод «расщипывания» нервной ткани, удалось выделить нервные клетки со всеми их отростками.
Нервная клетка с отходящими от нее отростками по предложению немецкого ученого В. Вальдеера (1891) получила название нейрона. Таким образом, нейрон является структурной единицей нервной ткани.
Другим структурным элементом нервной ткани считаются клетки глии — нейроглии. Будучи тесно связанными с нейронами, глиозные клетки, обладающие большим количеством отростков, представляют своеобразный опорный механизм, поддерживающий массу нейронов, а также выполняющий и ряд других функций — обменных, защитных и др.
Нейроны имеют различную форму, величину и характер отростков. Так, встречаются нейроны овальной формы, имеющие вид зерен, пирамидные, веретенообразные и др. Величина нейрона колеблется от 4 до 130 мкм. Цитоплазма нервной клетки {нейроплазмы) содержит обычные для всех типов клеток структурные части. В теле нейрона различают ядро и ядрышко, яв-
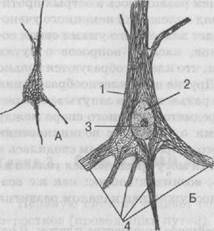
Рис. 19. Нервная клетка:
А. Общий вид; Б. 1 — тело клетки; 2 — ядро; 3 — ядрышко; 4 — отростки
ляющиеся наиболее важными составными элементами клетки (рис. 19). Вокруг ядра в цитоплазме после обработки метиленовой синью можно наблюдать своеобразные зерна синего цвета — хроматофильное вещество Ниссля (тельце Ниссля). Иногда эти зерна именуются тигроидным веществом или тигроидом (эти включения придают клетке своеобразную полосатость, напогтитя-ющую шкуру тигра). По>„. е импрегнации солями тяжелых металлов в нейроплазме выявляются тончайшие нити — нейрофибриллы. Электронно-микроскопическими
исследованиями показано, что нейрофибриллы состоят из пучков микротрубок различного диаметра. Эти структуры принимают участие в движении цитоплазмы (аксоплазматическом токе), а также в токе нейроплазмы в дендритах (рис. 20).
В цитоплазме нервной клетки можно встретить пигментные образования бурого или черного цвета — липофусцин и меланин.
От тела нейрона отходят отростки: короткие дендриты и длинные аксоны. В каждой клетке может быть несколько коротких отростков и один длинный. Отростки имеют своеобразные окончания. Так, короткие отростки заканчиваются мельчайшими ответвлениями, получившими название ши-пиков. Длинный отросток на конце разветвляется, образуя телодендрий. Тельца Ниссля обнаруживаются в дендритах, но не встречаются в аксонах. По дендритам к клетке поступают нервные импульсы. От нейрона импульсы распространяются по аксонам (рис. 21). Нейроны соединяются между собой при помощи своеобразных механизмов. Описано несколько форм межневронных соединений. Так, известный невро-гистолог С. Рамон-и-Кахал описал два типа таких соединений: 1) аксодендрический, при котором нити телодендрия соприкасаются с шипиками дендрита, что сопровождается выделением нейромедиатора ацетилхолина, вырабатываемого в
местах окончания отростков. Это соединение характерно для определенных групп клеток, находящихся в активном состоянии. Аксодендрический тип связей, по мнению С.А. Саркисова и Г.И. Полякова, преобладает в коре больших полушарий; 2) аксосоматический тип характеризуется образованием так называемой корзинки (по Гольджи), когда разветвления аксона оплетают все тело соседней клетки.
Так авторы нейронной теории представляли себе межклеточные соединения,образующие особые контакты, или синапсы (рис. 22). Однако механизм передачи возбуждения с одного нейрона на другой все же еще не представляется окончательно ясным. Имеется предположение о том (школа акад. К.М. Быкова), что механизм передачи возбуждения с одного нейрона на другой, в частности с афферентного на эфферентный1, обусловливается разностью электрических потенциалов, возникающих в области синапсов, что может быть в какой-то степени связано с образованием высокоактивных соединений типа упомянутого выше ацетилхолина.
По мнению В.А. Делова, образование в нервных клетках или в области синаптических окончаний ацетилхолина не исчерпывает всего цикла биохимических и физико-химических реакций, характеризующих деятельность центральной нервной системы, но является, по всей вероятности, обязательным
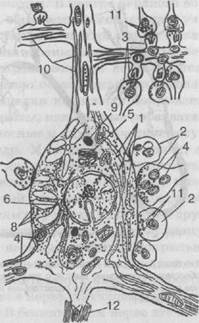
Рис. 20. Схема строения нервной клетки по данным электронной микроскопии (по Глезеру):
1 — канальцы эргастоплазмы (тигроид); 2 — аксосоматические контакты; 3 — аксо-дендрические контакты; 4— митохондрии; 5 — аппарат Гольджи; 6 — ядро клетки; 7 — ядрышко; 8 — микросомы; 9 — осмиофиль-ные тела; 10 — микротрубочки; 11 — си-наптические пузырьки; 12 — аксон с его миелиновой оболочкой
1 Афферентный — чувствительный нервный путь; эфферентный — двигательный нервный путь.
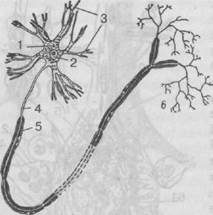
Рис. 21. Схематическое изображение нейрона:
1 — тело; 2 — ядро; 3 — дендриты; 4 — аксон; 5 — оболочки, образующие вместе с аксоном нервное волокно; 6 — конечные разветвления аксона (телодендрий)
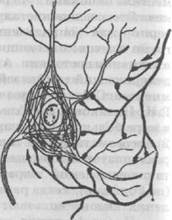
Рис. 22. Механизм синапса
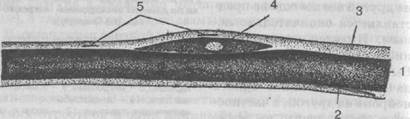
Рис. 23. Продольное сечение нервного волокна:
1 — осевой цилиндр; 2 — миелин; 3 — неврилемма; 4 — мякотная (покровная) клетка; 5 — ядро неврилеммы
звеном в цепи процессов, определяющих передачу возбуждения с нейрона на нейрон.
Роль так называемой нервной сети, состоящей из нейрофи-брилл, в процессах проведения нервных импульсов очень велика на низших уровнях развития животного мира. У млекопитающих, и особенно у человека, ее значение ограничено в связи с тенденцией к более тонкой дифференциации в структуре аппаратов, проводящих возбуждение.
Нервное волокно (рис. 23) представляет собой продолжение
отростков нейрона, в частности аксона. В центре нервного волокна проходит осевой цилиндр, образуемый скоплением пучков нейрофибрилл и представляющий центральный механизм, обеспечивающий проведение нервных импульсов. На некотором расстоянии от тела нейрона волокна покрываются двумя оболочками. Непосредственно осевой цилиндр обволакивает миелиновая оболочка. Миелин не сплошь покрывает осевой цилиндр, а образует перерывы, называемые перехватами Ранвье, куда впадают кровеносные и лимфатические сосуды, снабжающие осевой цилиндр. Миелиновая обкладка, в свою очередь, покрыта тонким, не имеющим структуры чехлом — неврилеммой, или шванновской оболочкой. Роль мие-линовой оболочки двоякая. С одной стороны, она предохраняет осевой цилиндр от всевозможных вредных влияний, с другой — ускоряет проведение нервных импульсов по нервному волокну. Различают мякотные нервные волокна, покрытые миелиновой оболочкой, и безмякотные (голые), входящие в состав симпатических нервов и обонятельных нитей. Скорость прохождения волны возбуждения в нерве, имеющем миелино-вую оболочку, от 60 до 120 м/с. В безмякотном нерве эта скорость меньше (от 1 до 30 м/с). Нервные волокна объединяются в нервные пучки и образуют периферические нервы. В крупных периферических нервах количество нервных волокон может доходить до нескольких тысяч. Это связано с тем, что этим нервам приходится снабжать громадное количество мышечных волокон, образующих скелетную мускулатуру.
Глия (невроглия). В состав нервных элементов, образующих нервную систему, включается еще один вид нервной ткани, известной под названием глии или невроглии. Эта ткань интимно связана с нейронами и их отростками, составляя по существу единую систему. По характеру клеточного строения нейроглия делится на микро- и макроглию. Для строения мак-роглии характерно наличие звездчатых клеток — астроцитов, обладающих большим количеством отростков, лучеобразно отходящих от тела клетки. Значение макроглии опорное: она как бы склеивает все элементы нервной системы, являясь своеобразным каркасом, поддерживающим массу нейронов. Мик-роглия состоит из клеток, выполняющих преимущественно трофические и защитные функции.
Нейронная теория углубила знания о характере строения нервной ткани. Однако следует помнить, что она создавалась в тот период, когда основные законы нервной деятельности, по-
строенные на рефлекторном принципе, еще не получили ведущей роли в неврологии. Идеи Р. Вирхова, представлявшего организм как механическую сумму органов и систем, имели главенствующее значение. Представители нейронной теории рассматривали нейрон не только как элемент структуры, но придавали ему значение физиологической единицы. Такое представление, естественно, приводило к неправильному пониманию целостной деятельности нервной системы, которая определялась как некая механическая сумма, складывающаяся из деятельности отдельных нейронов. Подобное мнение не могло удовлетворять современных сторонников нейронной теории.
Современная нейрофизиология определяет закономерности целостной деятельности коры больших полушарий, исходя из рефлекторного принципа. Отсюда направленность процессов возбуждения и торможения зависит от целого ряда различных влияний, а не только от деятельности отдельных нейронов. В этом случае большое значение получает то новое качество, которое создается в результате деятельности синапсов. Синапсы объединяют в новую качественную категорию отдельные нейроны. На основе этих связей и образуются физиологические механизмы, осуществляющие нервную деятельность, т.е. бесчисленные рефлексы головного и спинного мозга.
§2. Строение и функции головного мозга
Головной мозг как по строению, так и по своим функциям представляет исключительно сложный орган. Он является главным центром, в котором осуществляется взаимосвязь организма с внешней средой. В головной мозг через систему внешних рецепторов поступают сигналы из внешней среды. Внешний мир звуковых, световых, обонятельных, тактильных, вибрационно-кинестетических раздражителей влияет на наш мозг и особенно на его высшие отделы (кору), информируя о характере реальной действительности.
Помимо раздражителей из внешнего мира, головной мозг принимает импульсы из внутренней среды организма о состоянии внутренних органов. Таким образом, в высших отделах мозга осуществляется сложный анализ и синтез поступающих раздражений как из внешней, так и из внутренней среды, в результате чего образуются ответные распорядительные импульсы, регулирующие деятельность периферии.
Головной мозг живого человека имеет полужидкую консистенцию. Находясь в костном футляре (черепе), мозг, естественно, принимает форму черепа — шаровидную или несколько вытянутую (эллипсообразную) (рис. 24). Для удобства изучения вынутый из трупа мозг предварительно фиксируют в формалине, что сообщает ему большую плотность. В таком состоянии удобно производить разрезы с целью изучения особенностей его строения. Размеры головного мозга взрослого человека в среднем таковы: длина 170—180 мм, поперечник 140 мм, высота 125—135 мм (по Бунаку). Масса мозга в среднем у мужчин 1370—1380 г, у женщин 1250—1270 г. У детей школьного возраста масса мозга приближается к массе мозга взрослых. При изучении размеров и массы мозга обычно возникают вопросы о соотношении массы мозга с характером умственной одаренности. В специальной литературе приводятся цифры, характеризующие массу мозга выдающихся людей. Так, известно, что масса мозга писателя И.С. Тургенева составляла 2012 г, известного математика С. Ковалевской — 2000 г, Д. И. Менделеева — 1800 г, физика Лавуазье — 1700 г. В то же время имели место факты, когда масса мозга не менее выдающихся людей была значительно меньше. Так, мозг известного немецкого химика Либиха весил 1350 г. Известно, что масса мозга женщин в среднем ниже мужского на 120 — 150 г. Если исходить из ведущей роли массового показателя в одаренности человека, то можно прийти к абсурдным выводам о неполноценности женского мозга.
Какое же мнение может быть высказано по этому вопросу? Прежде всего, масса мозга вовсе не является единственным показателем, по которому можно судить об одаренности его обладателя, не говоря уже о том, что она может в значительной степени уменьшаться, в зависимости от условий пред-
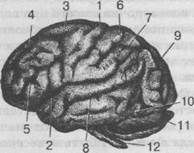
Рис. 24. Наружная поверхность головного мозга:
I — центральная борозда; 2 — боковая борозда; 3 — предцентральная борозда; 4 — верхняя лобная борозда; 5 — нижняя лобная борозда; 6 — зацентральная борозда; 7 — межтеменная борозда; 8 — верхняя височная борозда; 9 — затылочная теменная борозда (зарубка); 10 — предзатылочная зарубка;
I1 — мозжечок; 12 — продолговатый мозг
смертного периода (длительная и истощающая болезнь, старческая дряхлость), когда такой человек резко теряет массу всего тела. Имели место случаи большой массы мозга у отдельных лиц при относительно слабой умственной одаренности. Это может объясняться болезненным состоянием, выражающимся в патологическом увеличении серого вещества мозга (мегалоцефалия), или накоплением мозговой жидкости в желудочках мозга при водянке. Итак, изучение этого вопроса показало, что ни абсолютная, ни относительная (отношение массы головного мозга к массе тела) масса мозга не может служить показателем уровня интеллектуального развития.
Особенности физической конституции некоторых народностей (более рослые представители той или иной нации и менее рослые) также имеют значение в колебании массы мозга. Сюда же может быть отнесено и влияние половых различий. Так, например, сравнительно меньшая масса мозга у женщин не означает, что они обладают меньшими интеллектуальны ми возможностями, а закономерно обусловлена особенностями их скелета, мускулатуры, внутренних органов. Несравненно большее значение в развитии высоких умственных способностей имеет качество структуры корковых систем в целом или отдельных областей (слуховых, зрительных), заключающееся в особом развитии клеток, богатстве нервных связей. Однако важно иметь в виду не только структурные особенности, но и высокое качество тех нервных процессов, которые происходят в мозге человека, весь цикл условно-рефлекторной деятельности. Далее, следует помнить, что одаренность, талантливость не есть только результат механического, пассивного развертывания скрытых, заложенных от природы задатков, какого-то "золотого фонда". Многое в биологическом фонде опосредствовано социальными влияниями. Очень важен характер этих влияний, т.е. их сила, постоянство действия и т.п. Все это имеет самое прямое отношение к формированию типа нервной системы и определяет качество условно-рефлекторных связей, являющихся физиологической основой психики.
В этом смысле особое значение приобретают качество воспитания и обучения ребенка, приучение его к труду и последовательный, систематический труд. Гениальное произведение в науке или искусстве невозможно без напряженного труда. ".. .Наука, — говорил И.П. Павлов, — требует от человека всей
жизни, и если бы у нас было две жизни, то и их бы не хватило. Большого напряжения и великой страсти требует наука от человека". Когда мы читаем произведения классиков литературы, то поражаемся выразительности описания ими отдельных явлений природы, тонкости психологического анализа сложных переживаний отдельных героев. Если мы посмотрим подлинники их произведений, то увидим, что они пестрят бесчисленными переделками, поправками. Многие из знаменитых писателей или художников по несколько десятков раз переделывали страницы своих произведений или эскизы картин, добиваясь большей художественной правды. Все это есть результат тяжелого, напряженного труда, без которого не может быть гениального произведения ни в области искусства, ни в области науки.
Надо полагать, что в гениальности или одаренности того или иного человека, по-видимому, сочетаются два свойства. С одной стороны — это высокое качество биологического фонда (природных задатков), без которого невозможно развитие выдающихся способностей, с другой — это количество и качество той работы по самоусовершенствованию, по развитию своих способностей, которую производил данный человек, неуклонно стремясь к поставленной цели.
Головной мозг человека состоит из серого и белого вещества. Серое вещество составляют многочисленные клетки разнообразных форм и размеров. В коре больших полушарий насчитывается до 14 млрд клеток. Белое вещество состоит из волокон, имеющих также различное строение, причем одна группа волокон располагается горизонтально, а другие имеют радиальный (лучевой) тип расположения.
Мозг обильно снабжается кровью. Основными артериями, которые выполняют функцию мозгового кровоснабжения, являются внутренние сонные и позвоночные артерии. Последние поднимаются вдоль шейного отдела позвоночника и через большое затылочное отверстие проникают в полость черепа. Правая и левая позвоночные артерии соединяются между собой на основании мозга, образуя непарную мозговую артерию, ветви которой снабжают кровью задние отделы полушарий мозга, мозжечок, варолиев мост и продолговатый мозг. На основании головного мозга ветви задней мозговой артерии соединяются с передними ветвями сонной мозговой артерии, образуя кольцо (виллизиев круг) (рис. 25). От основного круга отходят передняя и средняя мозговая артерии, снабжаю-
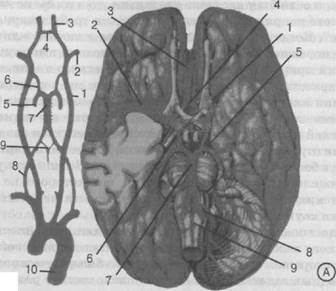
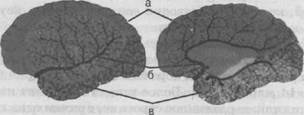
Рис. 25. Схема кровоснабжения головного мозга:
А. Схема артерий виллизиева круга: 1 — внутренняя сонная артерия; 2 — средняя мозговая артерия; 3 — передняя мозговая артерия; 4 — передняя спинальная артерия; 5 — задняя мозговая артерия; 6 — задняя соединительная артерия; 7 — основная артерия; 8 — позвоночная артерия; 9 — передняя спинальная артерия; 10 —аорта. Б. Корковые области кровоснабжения основными артериями головного мозга: а — область кровоснабжения передней мозговой артерией; в — область кровоснабжения задней мозговой артерией
щие все остальные части большого мозга, подкорковые образования, глубинные структуры мозга (рис. 26).
Левое полушарие обладает большим развитием сосудистой сети, следовательно, оно лучше снабжается кровью.
Некоторые авторы (Г. Грисбах, Л.В. Блуменау) связывают с этим обстоятельством и несколько больший вес левого полушария по сравнению с правым. У взрослого человека эта разница достигает 12 — 15 г. В левом полушарии расположены
особо важные сенсорные и моторные речевые центры. В правом полушарии этих центров не находили. Поэтому прежде считали некоторые зоны правого полушария немыми зонами. В настоящее время имеются данные о том, что нарушения модуляции голоса, монотонность речи, а также расстройство музыкальных способностей (амузия) могут возникать и при поражении правой лобной доли (Е.К. Сепп).
Отток венозной крови из мозга обеспечивается хорошо развитой системой вен. Малейшее затруднение в оттоке венозной крови вызывает повышение внутричерепного давления. Центральный венозный ствол, который собирает кровь из внутренних вен, называется веной Галена. Многочисленные вены головного мозга несут кровь в особые венозные пазухи, или синусы, которые заложены между листками твердой мозговой оболочки. Таких пазух имеется 21, из них 5 —
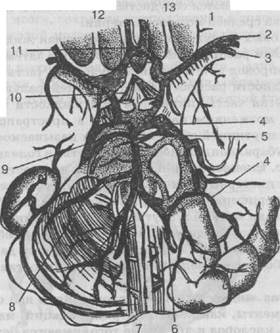
Рис. 26. Артерии головного мозга:
1 — средняя мозговая артерия; 2 — корковые ветви средней мозговой артерии; 3 — артерия сильвиевой борозды; 4 — задняя мозговая артерия: 5 — основная артерия; 6 — позвоночная артерия; 7 — передняя спинальная артерия; 8 — нижняя передняя мозжечковая артерия; 9 — верхняя мозжечковая артерия; 10 — задняя соединительная артерия; 11 — передняя соединительная артерия; 12 — внутренняя сонная артерия; 13 — передняя мозговая артерия
непарных. Синусы собирают кровь не только из мозга, но и из вен глазницы, уха и твердой мозговой оболочки. Из синусов венозная кровь поступает в большую яремную вену. Кроме артерий и вен, в мозге есть еще мельчайшие сосуды — капилляры, которые представляют конечные разветвления артерий. Капилляры делятся на прекапилляры, на уровне которых происходит газообмен крови, и посткапилляры, которые собираются в вены.
Мозг не имеет специальной лимфатической системы. Функцию этой системы в некоторой степени выполняет циркулирующая в желудочках мозга мозговая жидкость (ликвор).
Желудочки головного мозга представляют собой полости. Существуют боковые желудочки в каждом из полушарий мозга и по одному желудочку в межуточном и заднем мозге (четвертый желудочек, или ромбовидная ямка). В боковых желудочках располагаются сосудистые сплетения, к которым подходят ветви средней мозговой артерии.
Из боковых желудочков спинномозговая жидкость по узким каналам распространяется в третий, а затем через силь-виев водопровод в четвертый желудочек. Часть спинномозговой жидкости распространяется по центральному каналу вниз. Другая часть спинномозговой жидкости через отверстие под мозжечком направляется в пространство между мягкой и паутинной оболочками, так называемое подоболо-чечное (субарахноидальное) пространство в головном и спинном мозге. Скопление спинномозговой жидкости находится в верхней цистерне под мозжечком и в нижней цистерне, где имеются периферические нервы ("конский хвост"). Цистерна образована мягкой и паутинной оболочками. Желудочки головного мозга соединены не только между собой, но и с центральным каналом спинного мозга. Все это дает возможность свободного обращения ликвора в полостях головного мозга.
Мозговая жидкость в норме прозрачна, в ней содержатся такие элементы, как калий, натрий, кальций, магний, фосфор, азот, кислород и др. Кроме того, имеются белок, сахар, холестерин, а также небольшое количество лейкоцитов. Основное значение спинномозговой жидкости состоит в поддержании осмотического равновесия в нервной ткани, а также в защите нервной системы от всякого рода резких сотрясений, связанных с прыжками, бегом и т.д. Ряд авторов признает также питательное значение жидкости и ее участие в удале-
нии продуктов мозгового обмена. При патологии, в частности при воспалительных процессах, сообщающиеся отверстия могут закрываться и жидкость, не получая оттока, скапливается в замкнутой полости и давит на мозг, который в свою очередь оказывает давление на стенки черепа. Если это происходит у маленького ребенка, у которого швы между отдельными костями черепа еще непрочны, может произойти расхождение черепных костей и увеличение объема черепа. Такое состояние носит название водянки головного мозга (гидроцефалия).
Головной мозг заключен в специальные оболочки. Различают три мозговые оболочки — твердую, паутинную и мягкую.
Твердая мозговая оболочка состоит из плотной соединительной ткани, имеет волокнистое строение. Наружный ее слой служит надкостницей костей черепа, внутренний дает особые отростки, которые срастаются в определенных местах с веществом мозга, сохраняя его положение в полости черепа. Отростки твердой мозговой оболочки образуют венозные синусы. Паутинная оболочка покрывает сверху вещество мозга и состоит из рыхлой соединительной ткани, бедна кровеносными сосудами. Мягкая мозговая оболочка образуется из тонкой соединительной ткани, обильно снабженной кровеносны-
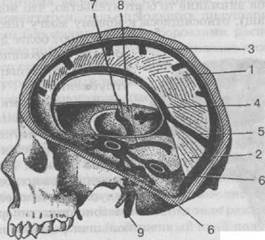
Рис. 27. Твердая мозговая оболочка и венные пазухи:
1 — большой серповидный отросток; 2 — палатка мозжечка; 3 — верхний серповидный синус; 4 — нижний серповидный синус; 5 — прямой синус; 6 — поперечный синус; 7 — верхний каменистый синус; 8 — нижний каменистый синус; 9 — яремная вена
ми сосудами, и сращена с верхними отделами коры головного мозга. Кровеносные сосуды из мягкой мозговой оболочки проникают в верхние отделы коры. Срастаясь с веществом мозга, мягкая мозговая оболочка повторяет рисунок коры головного мозга. Между паутинной и мягкой мозговыми оболочками образуется подоболочечное пространство, заполненное спинномозговой жидкостью.
Топография головного мозга
Для того чтобы лучше изучить особенности строения головного мозга, необходимо понять взаимоотношение составляющих его частей, т.е. его топографию. В этих целях вначале сделаем обзор общего рельефа мозга со стороны мозговых полушарий, далее рассечем мозг вертикально по средней линии, вдоль продольной щели и ознакомимся с особенностями внутреннего строения. Для ознакомления с нижней поверхностью (основанием) перевернем мозг основанием вверх и рассмотрим особенности структуры. Наконец, произведем горизонтально разрез, при котором увидим отделы мозга, которые на предыдущих срезах не были доступны обозрению.
При обзоре рельефа головного мозга (вертикальный срез) (рис. 28), и особенно со стороны внутренней поверхности, обращает на себя внимание то обстоятельство, что мозговые полушария (плащ), относящиеся к новому мозгу (неоэнцефалон),
покрывают более древние образования (палеоэнцефалон). Поверхность полушарий покрыта углублениями (бороздами); между бороздами находятся возвышения, называемые извилинами. Рисунок борозд и извилин (архитектоника) довольно сложен. Если сравнить строение поверхности больших полушарий человека и высших позвоночных, то сразу заметна большая разница в смысле усложнения архитектоник борозд и извилин у человека по сравнению с животными. Борозды, расположенные на поверхности полушарий, бывают двух ви-
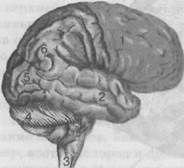
Рис. 28. Головной мозг (вид сбоку):
1 — лобная доля; 2 — височная доля; 3 — продолговатый мозг; 4 — мозжечок; 5 — затылочная доля; 6 — теменная доля
поз. Одни из них, глубокие и онтогенетически ранние, проходят через всю толщу стенки мозга и вдавливают последнюю в полость желудочков, образуя выступы. Другие представляют собой борозды, прорезывающие кору мозга, и ограничиваются только ею (И.Н. Филимонов). Глубокие борозды делят поверхность мозговых полушарий на определенные доли, являясь своеобразными естественными границами. Так, от середины верхнего края полушарий тянется глубокая борозда по направлению сверху вниз, это центральная, или роландова, борозда. Участок мозга, находящийся кпереди от центральной борозды, называется лобной долей. Кзади от центральной борозды лежит теменная доля, за исключением самого заднего отдела рельефа мозговых полушарий, образующих затылочную долю. На боковой поверхности полушарий располагается другая глубокая борозда, идущая в передне-заднем направлении, — боковая, или сильвиева, борозда. Часть мозга, находящаяся под сильвиевой бороздой, относится к височной доле. Таким образом, в каждом мозговом полушарии, а их два — правое и левое — выделяются четыре доли: лобная, теменная, затылочная и височная. Если широко раздвинуть края сильвиевой борозды, то можно увидеть выступ, испещренный бороздами и извилинами. Это дно сильвиевой борозды, образующее пятую долю мозга, которая называется островком. Помимо указанных борозд-ориентиров, рельеф больших полушарий покрыт более мелкими бороздами. Между бороздами располагаются извилины. Более подробно об этом будет сказано ниже. Для ознакомления со строением медиальной (внутренней) поверхности сделаем вертикальный разрез вдоль большой продольной расщелины, расчленив мозг на два полушария. На внутренней поверхности (рис. 29) можно отметить корковую часть, покрытую также бороздами и извилинами. В виде большой дуги тянется мозолистая борозда, окружающая мозолистое тело, поясная борозда и поясная извилина, шпорная борозда и др. Ниже коркового слоя на разрезе образование, имеющее форму полудуги. Это большая спайка мозга, или мозолистое тело. Оно соединяет два полушария. В мозолистом теле различают утолщение, тело, колено, клюв. Утолщенный край полушария под мозолистым телом, который проходит в виде дуги, образует так называемый свод. Ножки свода на основании мозга загибаются в височную долю и кончаются там бахромкой. Между мозолистым телом и сводом лежит треугольная пластинка, нося-Щая название прозрачной перегородки. Ниже, под сводом и
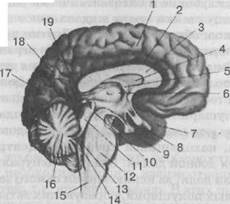
Рис. 29. Продольный разрез по средней линии головного мозга:
1 —лобная доля; 2 —поясная извилина; 3 —мозолистое тело; 4 —прозрачная перегородка; 5 — свод; 6 — зрительный бугор; 7 — височная доля; 8 — перекрест зрительных нервов: 9 — гипофиз; 10 — мозговой водопровод; 11 — пластинка крыши (четверохолмие); 12 — мост; 13 —верхний мозговой парус; 14 — четвертый желудочек головного мозга; 15 —продолговатый мозг; 16 — мозжечок; 17 — затылочная доля; 18 — теменная доля; 19 — эпифиз (шишковидное тело)
прозрачной перегородкой, лежит крупное образование, имеющее полуовальную форму. Это зрительный бугор, входящий в состав межуточного мозга. На поверхности зрительного бугра заметно небольшое возвышение — серая спайка, соединяющая зрительные бугры правого и левого полушарий. Внутренняя поверхность зрительных бугров образует стенки особой полос ти, называемой третьим мозговым желудочком, который представляет расширение подходящего сюда, желобка или канала, носящего название сильвиева водопровода. Нижний конец этого канала впадает в полость четвертого мозгового желудочка. Дном его является часть задней поверхности продолговатого мозга — ромбовидная ямка, а крышей.— тонкий складчатый листок, отходящий от мозжечка, — мозговой парус. Кзади от зрительного бугра имеется образование, представляющее собой вырост крыши межуточного мозга. Это шишковидная железа — эпифиз. Под зрительным бугром располагается нижний отдел межуточного мозга, называемый подбугорьем. Под-бугорье состоит из серого бугра, который переходит в воронку, на конце которой находится мозговой придаток — гипофиз, и двух образований, имеющих форму соска, — сосцевидных тел. Эпифиз и гипофиз относятся к железам внутренней секреции. Спереди к серому бугру прилегает перекрест зрительных нер-
вов — хиазма. Кпереди и кверху от хиазмы располагается передняя спайка мозга.
Зрительный бугор и подбугорье — гипоталамическая область — относятся к межуточному мозгу. С ним тесно граничит другой отдел головного мозга, который мы увидим на сагиттальном срезе. Это средний мозг. Последний состоит из четырех овальных бугорков, имеющих сложное слоистое строение и именуемых четверохолмием, и подходящих сюда двух толстых тяжей — ножек мозга, в которых проходят пути, связывающие кору с нижележащими отделами. Средний мозг включает в себя еще ряд образований, связанных с различными видами рецепций — слуховой, общей чувствительности, равновесия и др. На сагиттальном срезе средний мозг рассечен, и при обзоре мы видим только его половину — двухолмие и рассеченные вертикально ножки мозга. Средний мозг пересекает сильвиев канал, образующий выше расширение — третий мозговой желудочек. Ниже среднего мозга заметно выделяются нижние части ствола, которые представлены в виде удлиненного выступа или конуса, рассеченного по вертикали. Это варолиев мост и продолговатый мозг — отделы заднего мозга. Они в основном состоят из проводящих путей и заложенных в их толще ядер черепно-мозговых нервов. На этом срезе под затылочной долей мы увидим еще одно образование, которое располагается над продолговатым мозгом и варолие-вым мостом — мозжечок. Структура его внутреннего строения напоминает ветви дерева. Своеобразный рисунок строения мозжечка образован переплетениями серого и белого веществ, входящих в его состав. На внутреннем срезе доступны рассмотрению полости третьего и четвертого желудочков, соединенных между собой сильвиевым каналом.
Боковые желудочки, заложенные в толще полушарий, при данной проекции не видны, они соединяются с третьим желудочком при помощи монроевых отверстий. Четвертый желудочек при помощи отверстий Мажанди и Лушка соединен с под-оболочечными пространствами. Вся система полостей головного мозга соединена с полостью спинного мозга. В этой системе происходит циркуляция ликвора.
Для ознакомления с особенностями строения нижней поверхности мозга перевернем мозг основанием вверх (рис. 30). Здесь также можно увидеть ряд образований, с частью которых мы уже знакомились на вертикальном срезе. Из долей мозга на основании видна нижняя поверхность лобной, височ-
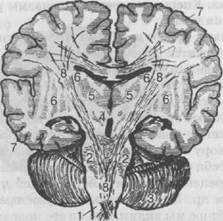
Рис. 30. Поперечный разрез мозга:
1 — продолговатый мозг; 2 — варолиев мост; 3 — мозжечок; 4 — средний мозг; несколько выше цифры располагается красное ядро; 5 — зрительные бугры; 6 — полосатое тело; 7 — кора больших полушарий; 8 — путь центробежных волокон от коры больших полушарий в спинной мозг (внизу виден перекрест волокон); 9 — волокна, соединяющие левое и правое полушария
ной и частично затылочной доли. На нижней поверхности лобной доли заметен ход обонятельных волокон, проходящих через обонятельную луковицу, погружающихся в толщу мозга и заканчивающихся в области крючковиднои извилины на внутренней поверхности полушарий (корковый центр обоняния). Несколько ниже заметно выделяется перекрест зрительных нервов — хиазма. Снаружи от хиазмы лежит передняя продырявленная пластинка — серая масса, пронизанная многочисленными сосудами и отростками мягкой оболочки. Это образование относится к обонятельной системе. Ниже хиазмы, в глубине полушарий, заметны два толстых тяжа, выходящих из толщи варолиева моста и направляющихся в кору полушарий. Это ножки мозга. Между ножками можно рассмотреть нижние отделы межуточного мозга — подбугорья, состоящего из серого бугра, двух сосцевидных тел, воронки, на которой помещается гипофиз. Ниже заметно выступают части мозгового ствола — варолиев мост и продолговатый мозг. Здесь хорошо видны отходящие из ядер ствола корешки тройничного нерва, выходящего из толщи варолиева моста. На границе между варо-лиевым мостом и продолговатым мозгом выходят корешки слухового, лицевого и отводящего нервов. Продолговатый мозг в поперечнике значительно уже варолиева моста. На пе-
редней поверхности продолговатого мозга обращают на себя внимание пучки продольно идущих волокон. Это пирамиды, в которых проходят двигательные пути. Несколько кнаружи от пирамид лежат парные овальные образования — оливы, волокна которых участвуют вместе с другими системами в образовании нижней ножки мозжечка. На основании хорошо видны отходящие от продолговатого мозга четыре пары черепно-мозговых нервов, ядра которых залегают в ромбовидной ямке.
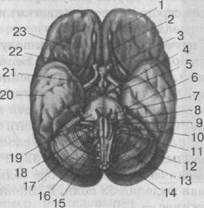
Рис. 31. Основание (нижняя поверхность) головного мозга:
1 — обонятельная луковица; 2 — обонятельный тракт; 3 — зрительный нерв; 4 — глазодвигательный нерв; 5 — блоковый нерв; 6 — тройничный нерв; 7 — отводящий нерв; 8 — лицевой нерв; Э — преддверно-улитковый нерв; 10 — языкоглоточный нерв; 11 — блуждающий нерв; 12 — добавочный нерв; 13 — мозжечок; 14 — затылочная доля; 15 — продолговатый мозг; 16 — подъязычный нерв; 17 — пирамида; 18 — олива; 19 — мост; 20 — височная доля; 21 — сосцевидное тело; 22 — гипофиз; 23 — лобная доля
Это языкоглоточный, блуждающий, добавочный и подъязычный нервы. Над стволом со стороны затылочной доли расположен мозжечок (рис. 31). Он состоит из двух полушарий и соединяющего их промежуточного звена — червя. Мозжечок связан с различными отделами головного мозга особыми путями, так называемыми ножками мозжечка. Таких ножек три пары: нижние ножки (веревчатые тела), осуществляющие вместе с оливарными волокнами и другими путями связь мозжечка с продолговатым и спинным мозгом; средние ножки, связывающие мозжечок с варолиевым мостом, и верхние ножки, связывающие мозжечок со средним мозгом.
Поперечные срезы головного мозга могут производиться на различных уровнях. Если произвести разрез через зрительный бугор, то можно, помимо ряда других образований, заметить
боковые желудочки, представляющие своеобразную щель с верхними и нижними разветвлениями — рога желудочка. На дне желудочков заметно сосудистое сплетение, продуцирующее мозговую жидкость.
Конечный, или большой, мозг
Конечный мозг в онтогенезе развивается из переднего мозгового пузыря и представлен двумя полушариями. Полушария мозга построены из серого вещества (клетки) и белого вещества (волокна или проводящие пути).
Микроскопическое изучение коры больших полушарий, начатое отечественным ученым профессором В.А. Бецем в 1869 г., Т. Мейнертом и другими, показало, что ее строение (архитектоника) неодинаково. Образующие кору клетки и волокна имеют в разных областях некоторые особенности структуры. В связи с этим кору делят на ряд участков (полей), характеризующихся толщиной и плотностью корковых слоев и особенностью строения составных элементов (рис. 32). Ряд авторов выделяли различное количество полей в коре больших полушарий. Так, К. Бродман выделял 52 поля, К. Экономо — 109, О. Фогт — 180, Г. Коскинас — 119.
В настоящее время в нейрогистологии принят шестислой-
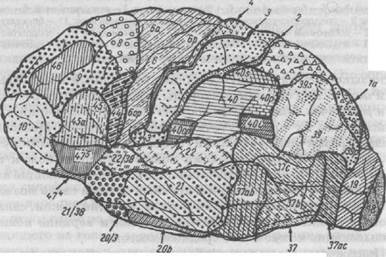
Рис. 32. Цитоархитектоническая карта коры мозга, наружная поверхность
ный тип строения коры больших полушарий (рис. 33). Выделяются следующие слои: первый — зональный — возникает рано, имеет светлую структуру, беден клетками; второй — наружный зернистый (преобладают клетки зерна); третий — слой пирамидных клеток; четвертый — внутренний зернистый (преобладают мелкие клетки-зерна); пятый слой — ганглионарный — встречаются большие пирамидные клетки; шестой — мультиформный, образован клетками треугольной и веретенообразной формы и чаще делится на два подслоя. Однако шестислойный тип строения выдержан не по всей коре. Так, например, исследования А. Каперса, И.Н. Филимонова и К. Экономо указывают на область передней центральной извилины, где зернистый слой вообще не представлен. Наоборот, в области шпорной борозды внутренний зернистый слой разделяется на три отдельных слоя. Аналогичные явления невыраженности ше-
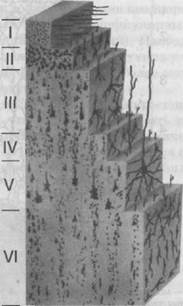
Рис. 33. Строение коры больших полушарий
Нейроны в коре головного мозга располагаются послойно. Каждый слой, кроме первого (I), образованного отростками нервных клеток, составлен в основном из нейронов однотипной конфигурации. Во втором (II) слое располагаются в основном клетки-зерна, в третьем (III) — пирамидные нейроны, в четвертом (IV) слое — звездчатые, в пятом (V) — сконцентрированы крупные пирамидные двигательные нейроны, в шестом (VI) — веретенообразные
стислойного строения наблюдались в области крючковидной извилины (корковый центр обоняния). Примерно */\% часть коры не имеет строго выдержанного шестислойного строения. Эта часть коры относится к так называемой старой коре (аллокортекс), которая является переходной формацией от коры высших животных к человеку. Новая кора (неокортекс) у человека составляет 96% всей поверхности больших полушарий. Большей частью она характеризуется шестислойным строением. Волокна коры представляют собой отростки нейронов. Обычно их подразделяют на Две группы. Одни имеют вертикальное (лучистое) направле-

Рис. 34. Цитоархитектоническая и миелоархитектоническая схема коры головного мозга:
1 — молекулярный слой; 2 — наружный зернистый слой; 3 — слой малых и средней величины пирамидальных клеток; 4 — внутренний зернистый слой; 5 — слой больших пирамидальных клеток; 6 — слой полиморфных клеток; 7 — белое вещество
ние, другие тянутся горизонтально. Среди горизонтальных волокон выделяют различные типы, так называемые полоски. На рис. 34 показаны особенности строения клеток и волокон в различных слоях коры. Функциональное значение отдельных слоев еще окончательно не выяснено. Есть основание считать, что верхние слои выполняют ассоциативные (связующие) функции, четвертый (зернистый) слой несет преимущественно функции рецепции, пятый и шестой слои имеют отношение к двигательным актам.
В коре головного мозга выделяют лобную, теменную, височную и затылочную доли. Рассмотрим их характерные особенности.
Лобная долязанимает передний фасад мозгового полушария и представляет собой участок коры, отграниченный от теменной доли (сзади) роландовои бороздой и от височной доли (снизу) сильвиевой бороздой.
Лобная доля покрыта бороздами и извилинами, среди которых выделяют прецентральную борозду, идущую параллельно роландовои. Между ними располагается передняя центральная извилина. В переднем отделе лобной области выделяются верхняя и нижняя лобные борозды, между которыми распола-
гаются верхняя, средняя и нижняя лобные извилины. По своему строению и функции нижняя лобная извилина неоднородна и подразделяется на три отдела: верхний (оперкулярный), средний (триангулярный) и нижний (орбитальный). Наибольшее значение эта область коры головного мозга получила у человека, так как связана с речевой функцией. Наиболее молодой областью является лобный конус.
Кора лобной доли самая толстая (2,5 — 4,5 мм). Микроскопическое строение ее неоднородно, что позволяет выделить ряд корковых полей, имеющих различную функцию. Первичное моторное поле (рис. 32, 35) (поле 4), от которого начинается пирамидный путь, располагается в передней центральной извили-
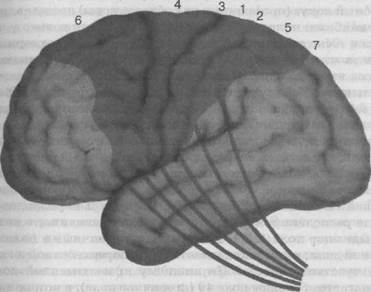
Рис. 35. Корковое представительство пирамидного пути:
4 — первичное двигательное поле, 1,2,3,5,6,7 — адверсивные поля
не. Кпереди от поля располагается поле б (вторичное моторное поле), обеспечивающее переключение из одного положения в Другое. Оба поля работают в тесном содружестве. Нижние отделы полей 4 и 6 иннервируют мышцы лица, артикуляции, глотки. Средние отделы обеспечивают движение верхних конечностей и особенно мелкую моторику пальцев рук. Верхние отделы иннервируют нижние конечности. Установлено, что поле 4, осо-
бенно в нижней его части, сдвинуто в роландову борозду, а на наружной поверхности занимает область нижней лобной извилины, имеющей отношение к речи (поля 44 и 45). Рядом с отделами, иннервирующими артикуляционную мускулатуру, располагается участок коры, обеспечивающий общую и мелкую моторику рук. Речь и труд — основа человеческой деятельности, тесно связаны между собой. Верхние отделы передней центральной извилины, посылающие импульсы к нижним конечностям, переходят на внутреннюю поверхность коры головного мозга между двумя полушариями. В средней лобной извилине (соответственно участку иннервации лицевой мускулатуры поля 4 и 6) располагается поле 8, обеспечивающее содружественные движения головы и глаз при повороте в сторону.
Лобный конус (префронтальная область коры) представляет собой наиболее молодое образование, имеющееся только у человека и тесно связанное со всеми отделами коры, подкорки и ствола мозга. Лобный конус является пусковым механизмом для всех видов высшей психической и эмоционально-волевой деятельности. Базируясь на тесном контакте с речевой областью, лобный конус (поле 10) формирует произвольную деятельность, все высшие психические функции человека.
Теменная доля занимает верхнебоковой фасад полушария. Границей ее является спереди роландова борозда, снизу силь-виева борозда, сзади непостоянная передняя затылочная борозда. Кора теменной области обладает по сравнению с лобной меньшей толщиной — 2,5—1,5 мм. Параллельно роландовой борозде располагается задняя центральная извилина, в которой выделяют поля 3,2,1, являющиеся первичными полями теменной доли; в них заканчивается путь поверхностной и глубокой чувствительности. По нижнему краю теменной доли располагается большое поле 40 (вторичное поле), в котором на основании восприятия поверхностной и глубокой чувствительности (кинестезии) фиксируются заученные движения — пра-ксис. Для артикуляционной мускулатуры это нижние отделы поля 40 — оральный праксис. Третичное поле теменной области (поле 39) располагается на стыке теменной, затылочной и височной областей и выполняет сложную функцию формирования зрительно-пространственного синтеза (конструктивный праксис) — ориентация в пространстве и в схеме тела. Становление этого отдела теменной области происходит значительно позже и в тесном сотрудничестве со зрительной и слуховой долями мозга.
Затылочная доля занимает задний отдел мозговых полушарий, тесно связана с теменной и височной долями мозга и не имеет выраженных границ. Это наиболее старое образование, толщина которого 1,5—2,5 мм. Микроскопическое строение коры неодинаково, в связи с чем выделяют всего три поля — 17,18,19. Со стороны пито- и миелоархитектоники особенно развит четвертый (зернистый) слой с преобладающим горизонтальным типом волокон. Первичным полем затылочной области является поле 17, в котором заканчивается зрительный путь. Так как данное поле является наиболее старым образованием, оно полностью сдвинуто на внутреннюю поверхность мозга. В поле 17 фиксируются не элементарные формы зрительного анализа и синтеза, а комплексные. Вторичное поле 18 надстроилось над первичным и выполняет в содружестве с другими отделами функцию зрительного восприятия. Наиболее сложным по структуре является третичное поле 19, расположенное на наружной поверхности коры (зона перекрытия с височной и теменной областью) и тесно связанное с речью.
Височная доля мозга занимает нижнебоковой фасад полушария. Этот участок коры отграничен от лобной и теменной долей сильвиевой бороздой. Граница с затылочной долей условная. На наружной поверхности височной доли располагаются три височные борозды : верхняя, средняя и нижняя. Между ними находятся соответствующие височные извилины. В височной области выделяется поперечная извилина Гешля, в которой заканчивается слуховой путь. Будучи наиболее старой, извилина Гешля полностью сдвинута в сильвиеву борозду. В ней располагаются поля 41 и 42 — первичные (проекционные) поля слуховой зоны. На наружной поверхности височной доли располагается верхневисочная извилина (поле 22), в которой преобладают ассоциативные слои клеток, связывающие височную долю мозга с другими отделами коры, в частности с затылочной, теменной и моторной (нижнепремоторными областями коры). Средние отделы височной области — поле 21, оно является третичным полем, имеется только у человека и связано с восприятием и удержанием в памяти словосочетаний (лексико-грамматических структур).
На границе височной и затылочной областей коры головного мозга формируется поле 37, в котором фиксируются комплексные раздражения, приходящие в слуховую и зрительную области. Эти связи начинают формироваться очень рано (в доречевом периоде), обеспечивая возможность восприятия речи и оречевление окружающих предметов. Эта об-
ласть коры тесно связана с премоторными отделами лобной области коры, обеспечивая возможность речевого развития ребенка, а также повороты головы и глаз на звук (в сторону звучащего тела).
Лимбическая область коры головного мозга (лимбическая, или краевая, кора) является древним образованием, располагается на нижней и внутренней поверхности полушарий. Она сочетается с миндалевидным ядром из группы подкорковых образований, обонятельным нервом, участками лобной, теменной и височной долей коры больших полушарий мозга, а также с под-бугровой областью и ретикулярной формацией, объединяясь в лимбико-ретикулярную систему. Это сложное многоуровневое образование обеспечивает регуляцию тонуса коры, сложные разноплановые функции внутренних органов, мотивационно-эмоциональные реакции и сложные инстинкты, поддерживает связь с внешним миром. На ранних этапах филогенеза эта область сформировалась в тесной связи с обонятельным нервом, выполнявшим сложные функции и имевшим большое значение в жизни живых существ (обонятельный мозг). В жизни человека обоняние потеряло свою первоначальную значимость, но сохранилось как воздействие на эмоциональную сферу, состояние соматических и висцеральных функций.
Локализация функций в коре головного мозга
Исследование структуры переднего мозга, естественно, не могло быть оторвано от изучения функционального значения отдельных областей коры. Проблема локализации функций в коре больших полушарий давно являлась одним из сложнейших разделов в научном естествознании. Во второй половине XIX в. было открыто клеточное строение организма, получила полное подтверждение эволюционная теория в известных трудах Ч. Дарвина, Д.И. Менделеевым была открыта периодическая таблица химических элементов, а наука о мозге не сделала сколько-нибудь существенных сдвигов вперед. Характеризуя именно этот период, И.П. Павлов говорил: "Неудержимый со времен Галилея ход естествознания заметно приостанавливается перед высшим отделом мозга, или, вообще говоря, перед органом сложнейших отношений животных к внешнему миру"1. Спорным являлся вопрос об особенностях работы мозга.
1ПавловИ.П. Поли. собр. соч. — М., Изд-во АН СССР, 1951. — Т. 3. —С.113.
Как работает мозг? Имеются ли в его строении отдельные пункты, центры, избирательно управляющие отдельными функциями, или весь мозг в целом принимает участие в регуляции различных функций, и особенно психических?
Еще в 1825 г. французский ученый Ж. Галль предположил, что вся мозговая кора покрыта многочисленными центрами, которые управляют различными психическими способностями. Хорошее развитие отдельных способностей (например, математических) обусловливается, по Галлю, усиленным развитием корковой массы в определенном центре. Это приводит к выбуханию, неровностям на поверхности черепа, возникающим в результате давления определенных участков коры на внутренние стенки костей черепа (отсюда выражение — "математические шишки"). Учение Галля (френология) не было подтверждено соответствующими научными исследованиями и вскоре было отвергнуто. Противоположные взгляды по вопросу о локализации функций высказывали М. Флуранс, Ф. Мажанди и др., которые считали, что кора функционирует как единое целое в каждом акте и специальных локальных центров нет. Подобные взгляды, утверждавшие эквипотенциальное (равнозначное) значение всех отделов коры, также не были правильными.
Последующие исследования установили особое значение моторного центра речи, опровергли мнение противников локализации. Выводы П.Брока строились на сопоставлении клинических наблюдений над больными афазией с последующими патологоанатомическими данными посмертного вскрытия мозга. Это открытие послужило началом развития учения о локализации функций. Дальнейшие классические эксперименты, проведенные по методу раздражения и экстирпации О. Фричем и Е. Гитцигом, а позже Г. Мунком, внесли много ценного в учение о локализации функций в коре больших полушарий. Эти исследователи, раздражая электрическим током, а также удаляя или разрушая определенные участки коры, наблюдали, какие изменения происходят в дальнейшем поведении животного. В результате таких экспериментов ими было установлено наличие двигательных центров в коре лобной доли. Последующие опыты Г. Мунка, Д. Ферриери, позже В.М. Бехтерева установили наличие в коре не только двигательных, но и слуховых, зрительных, обонятельных, вкусовых центров общей чувствительности. В этот период П. Флек-сиг отмечает значение лобных и нижнетеменных долей в пси-
хических процессах. К. Вернике указал на связь задней части верхней височной извилины с рецептивной функцией речи (сенсорный центр речи). Г. Липман выявил роль теменной доли в функциях праксиса1.
Таким образом, постепенно было установлено, что кора мозга не является эквипотенциальной (равнозначной) в отношении локализации отдельных функций. Исследования того времени показывали, что в лобной доле находится двигательный центр, управляющий движениями скелетной мускулатуры и движениями речевых органов, а также локализованы центры психических процессов ("верховные" центры). Теменная доля тесно связывалась с регуляцией различных процессов общей чувствительности. В височной доле определяли центр слуха и центр понимания речи (сенсорный центр), в затылочной — центр зрения, в крючковидной извилине, на внутренней поверхности мозгового полушария, — центр обоняния; спорной являлась локализация вкусового центра, местонахождение которого относили к оперкулярной области лобной доли.
Несмотря на правильную тенденцию, наметившуюся в исследованиях по изучению функционального значения отдельных областей мозга в общей целостной деятельности коры, все лее существовал ряд противоречий, создавались различные направления, причем одна группа ученых больше держалась локалистического направления, придавая различным участкам коры строго определенную функциональную значимость. В этом смысле они описывали и различные центры психических процессов (центры внимания, памяти-, воли, характера и т.д.), приурочивая их к определенным участкам мозга. В подобном аспекте рассмотрения, как это указывает А.Г. Иванов-Смоленский, действовал принцип анатомо-психологических корреляций, т.е. тому или иному анатомическому участку коры приписывалось выполнение определенных психических функций, при этом совершенно исключалось физиологическое звено.
Особенно узким локализационизмом отличались работы немецкого ученого К. Клейста.
Другая группа исследователей стояла на несколько иных позициях, придавая большее значение целостным реакциям
*Праксис — заученные движения, приобретаемые в процессе личного опыта.
коры, нежели локальным признакам. Позже это нашло отражение в работах К. Лешли, который считал, что локальный признак имеет второстепенное значение, а расстройство функции связано с количественным моментом (размеры пораженной массы мозга). К. Гольдштейн подчеркивал важность тех перестроек, которые возникают во всей остальной коре при поражении отдельного участка, что резко изменяет характер корковых функций.
Таким образом, рассмотрение в историческом аспекте развития учения о локализации функций показывает, что в основном существовали два направления, явно противоречащие ДРУГ Другу.
В чем причины неудовлетворительного решения вопроса о локализации функций в коре больших полушарий на прошлых этапах развития науки? Где источник тех постоянных противоречий, которые возникали у представителей разных направлений? Не говоря уже о том, что этот вопрос вообще представлял большую сложность, существовали определенные причины, которые задерживали успешное разрешение этой проблемы. Прежде всего, в работах старых авторов отсутствовал определенный методологический принцип, на основе которого можно было бы строго научно трактовать наблюдаемые факты. Отсутствие диалектического подхода приводило к неразрешимым противоречиям при постановке указанной проблемы. Рассмотрение этой проблемы на основе диалектического метода, в котором подчеркиваются взаимосвязь и взаимообусловленность различных процессов, протекающих в организме, в корне устраняет эти кажущиеся противоречия. Локализованная работа отдельных участков мозга вовсе не должна противостоять целостным реакциям коры, это лишь противоположные стороны единого процесса мозговой деятельности. Второй причиной, затруднявшей разрешение этой проблемы, являлось слабое развитие в тот период научных исследований в области физиологии мозга.
Исключительное значение приобретают работы русских Ученых И.М. Сеченова, открывшего рефлекторный принцип в работе головного мозга, и И.П. Павлова, установившего определенные закономерности протекания нервных процессов в коре больших полушарий в норме и патологии. Эти великие открытия выдвинули русских ученых в области естествознания в передовые ряды деятелей мировой науки.
Созданное И.П. Павловым учение о физиологической деятельности мозга углубило работу невропатологов и психиатров в клинике. В частности, благодаря трудам И.П. Павлова проблема локализации функций в коре больших полушарий получила новое освещение. Был сделан крупный шаг вперед на пути развития этой проблемы.
В настоящее время работами А.Р. Лурия и его школы доказано, что каждая область коры головного мозга выполняет определенную функцию и имеет особенности анатомического строения и физиологического характера.
Современное представление о локализации функцийв коре головного мозгасвязано с работами А.Р. Лурия, Г.И. Полякова и их школы.
Сопоставляя анатомические, физиологические и клинические наблюдения о значении долей мозга и выделенных в них полей, наука доказала их различия и раскрыла их особенности. Так, в каждой доле имеются первичные, вторичные и третичные поля. Особенностью первичных полей является структурная и функциональная организация сома-тотопической проекции, при которой отдельные точки периферии (кожной поверхности, скелетной мускулатуры, сетчатки глаза, слуховых клеток) проецируются в строго ограниченные соответствующие участки первичных полей коры головного мозга. Первичные поля непосредственно связаны с периферией, в них либо начинается путь (пирамидный путь от лобной области коры), либо заканчивается (зрительный путь в затылочной доле, слуховой путь в височной доле, путь общей и глубокой чувствительности — в теменной доле мозга).
В передней и задней центральной извилине на наружной поверхности мозга находится большой участок для представительства лицевой и артикуляционной мускулатуры, несколько выше (средняя треть извилины) располагаются участки мозговой коры, представляющие интересы руки и особенно пальцев рук. На самой вершине извилины на переходе на внутреннюю поверхность мозга располагаются представительства нижних конечностей. Такое расположение двигательных и чувствительных отделов в передней и задней центральной извилине указывает на значимость для человека артикуляционной моторики и ручной умелости. Нижние конечности, менее значимые для человека, сдвинуты на внутреннюю поверхность коры (рис. 36).
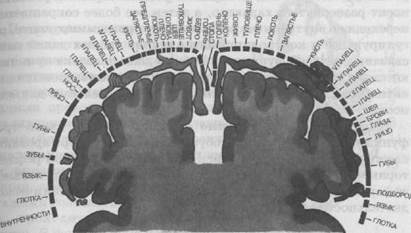
Рис. 36. Отделы поверхностей тела в проекционных зонах мозговой коры:
слева — корковая проекция чувствительности, справа — корковая проекция двигательной системы. Относительные размеры органов отражают ту площадь коры головного мозга (обозначено черной штриховой линией), с которой могут быть вызваны соответствующие ощущения и движения.
Вторичные поля, надстраиваясь над первичными, выполняют сложную интегративную функцию в организации двигательных актов, таких, как повороты головы и глаз, головы и всего тела, хватательные движения руки, переключения в общей и артикуляционной мускулатуре.
Наибольшее значение в речевой функции имеют третичные поля доминантного полушария мозга. Получая мощные импульсы от всех отделов коры головного мозга и нижележащих отделов, третичные поля коры играют важную роль в регуляции состояний активности, приводя их в соответствие с формулируемыми замыслами и намерениями. Наряду с регуляцией состояния активности, префронтальные области мозга принимают участие в формировании поведения человека. Темп роста префронтальных площадей мозга усиливается к 3—4 годам и к • 8 годам, что соответствует критическим периодам развития нервной системы.
Нижние отделы лобной коры связаны с формированием лексических и грамматических структур. Регуляция сознательной деятельности человека совершается при участии речи, психические процессы формируются и протекают на основе
речевой деятельности, которая на ранних ступенях развития носит развернутый характер, а затем все более сокращается, переходя во внутреннюю речь, поэтому программирующее, регулирующее и контролирующее влияние лобных долей тесно связано с речевой функцией. Лобные доли человека принимают самое непосредственное участие в экстренном повышении активности, сопровождающем всякую сознательную деятельность.
В заключение необходимо отметить тесную связь трех функциональных блоков, обусловливающих формирование сложной саморегулирующейся системы, называемой рефлекторным кольцом. Каждое ее звено включает афферентные и эфферентные компоненты и определяет характер психической деятельности (А.Р. Лурия, А.Н. Леонтьев).
Подкорковая область
Подкорковые образования — это скопление серого вещества, ближайшего к коре головного мозга. По времени возникновения, структуре и значению подкорковые образования различны. Здесь выделяют: хвостатое ядро, чечевичное ядро, ограду и миндалевидное ядро (рис. 37). Хвостатое ядро сформировалось из переднего пузыря и по своему происхождению ближе к коре головного мозга. Чечевидное ядро подразделяется на скорлупу и бледный шар. Близкие по своей структуре скорлупа и хвостатое ядро, а также более поздние образования составили ядро, называемое стриатум (полоса* тое тело). Бледный шар (паллидум) — более старое образование, антагонист полосатого тела. Полосатое тело и бледный шар образуют стрио-палл ид арную систему. Миндалевидное ядро тесно связано с лимбической областью. Значение ограды неясно.
Строение подкорковых узлов достаточно сложно. Так, для полосатого тела характерно наличие как крупных, так и мелких полигональных клеток, отличающихся хроматофильной цитоплазмой и большим количеством дендритов. В структуре бледного шара преобладают треугольные и веретенообразные клетки, много волокнистых образований.
Подкорковые узлы связаны между собой, а также с корой, межуточным и средним мозгом. Связь подкорковых узлов с корой осуществляется через зрительный бугор и его проводники. Некоторые исследователи признают наличие непосредст-
венной связи коры с подкорковыми узлами.
Подкорковые узлы окружены белым веществом, носящим своеобразное название — сумка. Различают внутреннюю, наружную и внешнюю сумки. В сумках проходят различные проводящие пути, осуществляющие связь коры с нижележащими областями и непосредственно с подкорковыми узлами. В частности, пирамидный путь, осуществляющий связь коры с различными этажами головного и спинного мозга, проходит через внутреннюю сумку. В функциональном отношении подкорковые узлы, являясь базой непроизвольных движений, принимают участие в сложных двигательных актах. Они являются также базой сложных безусловных рефлексов — пищевого, оборонительного, ориентировочного, полового и др., которые составляют фундамент высшей нервной деят
Дата добавления: 2019-07-26; просмотров: 863;