Действие инсулина на метаболические процессы
| процесс | печень является наиболее важным объектом при нагрузке глюкозой транспорт глюкозы свободный по градиенту концентрации | мышцы инсулин увеличивает проницаемость покоящейся мышцы для глюкозы. В работающей мышце транспорт глюкозы независим от инсулина. | жировая ткань инсулин увеличивает транспорт глюкозы в клетку. Конечными продуктами являются a-глицерофосфат и жирные кислоты |
| синтез гликогенаÝ | активирует гликогенсинтетазу | активирует гликогенсинтетазу | незначительно активирует гликогенсинтетазу |
| Гликоген олиз ß | снижает активность фосфорилазы. Для этого эффекта нужны небольшие количества инсулина. Выход - свободная глюкоза | снижает активность фосфорилазы. Выход - пируват и лактат | |
| гликолизÝ | повышает активность глюкокиназы и фосфофруктокиназы | повышает активность глюкокиназы и фосфофруктокиназы | такой же, но незначительный эффект |
| глюконео-генезß | снижает активность фосфоэнолпируват-карбоксикиназы. Для этого эффекта нужны большие количества инсулина. | ||
| липогенезÝ | стимулирует ацетил-СоА-карбоксилазу | ||
| липолизß | ингибирует гормончувствительную липазу липолиз снижается при увеличении количества субстрата - глицерина | ||
| обмен белков | увеличивает поглощение большинства аминокислот, стимулирует синтез белка, тормозит распад белков и снижает окисление аминокислот |
Снижение концентрации глюкозы в крови стимулирует секрецию глюкагона. Под влиянием глюкагона происходит распад гликогена в печени до глюкозы. В результате этого повышается содержание глюкозы в крови.
ГЛЮКАГОН - пептидный гормон, участвующий в поддержании углеводного обмена; является физиологическим антагонистом инсулина, а также стимулятором его секреции. Глюкагон секретируется альфа-клетками островков поджелудочной железы животных и человека. Помимо альфа-клеток поджелудочной железы, глюкагон секретируется аргирофильными клетками слизистой оболочки двенадцатиперстной кишки.
Биологическое действие глюкагона в огрганизме происходит в трех основных направлениях: а) активное участие в процессе гомеостаза глюкозы; б) стимуляция секреции инсулина и в) липолитическое действие. Гипергликемический эффект глюкагона при введении животным обусловлен его воздействием на процесс распада гликогена в печени. Это связано со стимуляцией глюкагоном перехода неактивной формы фосфорилазы В в активную фосфорилазу А, которая вызывает распад гликогена до глюкозо-6-фосфата. Гликогенолитическое действие глюкагона наступает при концентрации его 1 ммкг/мл.
В норме глюкагон предотвращает чрезмерное снижение глюкозы в крови, которое может произойти при усилении секреции инсулина. При снижении уровня сахара в крови выделение глюкагона увеличивается, что приводит к восстановлению содержания глюкозы до исходного уровня. Глюкагон оказывает значительно более выраженное гликогенолитическое действие, чем адреналин; в отличие от адреналина, также усиливающего распад гликогена в печени, глюкагон не влияет на распад гликогена в мышцах. Установлено, что физиологическое действие глюкагона двояко: как антагониста и как стимулятора секреции инсулина.
Глюкагон - липолитический гормон, мобилизующий жирные кислоты из жировой ткани. При этом также проявляется его контринсулярный эффект, поскольку инсулин способствует липогенезу. Механизм липолитического действия глюкагона проявляется через систему аденилатциклазы, активирующей ферменты липолиза.
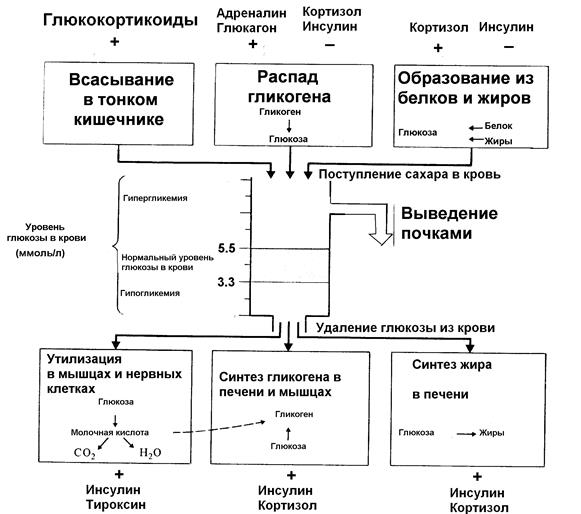
Рисунок 18. Гормональная регуляция уровня глюкозы в крови
В экспериментах глюкагон способствует секреции тиреокальциотонина и тем самым вызывает снижение содержания кальция в крови; Глюкагон оказывает положительное хронотропное действие на синусовый ритм, положительное инотропное действие на миокард, ускоряет предсердно-желудочковую проводимость и несколько уменьшает периферическое сопротивление. Это воздействие происходит независимо от действия катехоламинов и обусловлено активированием аденилатциклазы и гликогенолиза в мышце сердца.
В состоянии покоя клетки мозгового слоя надпочечников постоянно секретируют небольшие количества адренаналина и норадреналина. Под влиянием внешнего или внутреннего стрессирующего фактора, например в ситуациях, требующих большого физического или умственного напряжения, а также при инфекции и травме, резко повышаются секреция адреналина и норадреналина. Катехоламины стимулируют гликогенолиз в печени и мышцах. Увеличение синтеза цАМФ под влиянием катехоламинов и в большей степени адреналина активирует фосфорилазу печени, распад гликогена и образование большого количества свободной глюкозы. При этом увеличиваются поглощение кислорода, затраты энергии в связи с усилением сердечной деятельности, повышением мышечного тонуса и окислением молочной кислоты в печени.
Глюкокортикоиды вызывают повышение содержания глюкозы в плазме крови (гипергликемия). Этот эффект обусловлен стимулированием процессов глюконеогенеза в печени, т.е. образования глюкозы из аминокислот и жирных кислот. Глюкокортикоиды угнетают активность фермента гексокиназы, что ведет к уменьшению утилизации глюкозы тканями.
Соматотропный гормон при длительном повышении концентрации становится контринсулярным. Гормон роста увеличивает выход глюкозы в печеночные вены, усиливает глюконеогенез, уменьшает поглощение глюкозы на периферии, а также усиливает липолиз, в результате чего в крови повышается концентрация свободных жирных кислот, которые подавляют действие инсулина на мембранный транспорт глюкозы.
Жировой обмен
Жиры являются одним из основных источников энергии: 40-50% энергопродукции организма обеспечивается триглицеридами (триацилглицерины), на долю которых приходится более 95% всех липидов. В организме нормального человека с массой 70 кг на долю жировой ткани приходится 12 кг (110 000 ккал). Наряду с этим в крови определяется дополнительно 3 г триглицеридов (30 ккал), 0,3 г свободных жирных кислот (3 ккал) и 0,2 г кетоновых тел (0,8ккал).
Липогенез – процесс синтеза жирных кислот, интенсивно протекающий в печени и жировой ткани. При эстерификации жирных кислот с глицерином образуются триглицериды. В печени и жировой ткани инсулин увеличивает синтез жирных кислот и триглицеридов. Липолиз – процесс гидролиза липидов с образованием неэстерифицированных жирных кислот и глицерина – катализируется внутриклеточной гормональнозависимой липазой (триацилглицериновая липаза), которая воздействует на триглицериды и стимулируется цАМФ. Липаза лимитирует скорость липолиза, а образование цАМФ под влиянием аденилатциклазы находится в свою очередь под контролем различных гормонов.
Мембрана адипоцитов содержит рецепторы, взаимодействующие с гормонами, обладающими липолитическими свойствами (катехоламины, АКТГ, СТГ), и рецепторы к инсулину. В результате действия липолитических гормонов повышается активность аденилатциклазы, увеличивается образование цАМФ, активизируются гормончувствительная липаза и липолиз. Взаимодействие инсулина с соответствующими рецепторами, наоборот, приводит к угнетению аденилатциклазы, снижению концентрации цАМФ и торможению липолиза. Липолиз увеличивается во время голодания, при продолжительной работе, охлаждении, стрессе. Липолитическое действие катехоламинов (адреналина, норадреналина) и глюкагона осуществляется путем активации аденилатциклазы. С физиологической точки зрения, роль норадреналина в процессе липолиза представляется более важной, чем адреналина. Он образуется в адренергических нервных окончаниях в жировой ткани и обеспечивает мобилизацию жирных кислот.
Гормон роста оказывает мощное липолитическое действие, которое отличается от действия катехоламинов. Он вызывает увеличение концентрации свободных жирных кислот в плазме через 2-3 часа. Это действие, отмечающееся при введении даже небольших доз СТГ, по-видимому, связано с торможением процесса реэстерификации свободных жирных кислот. Тем не менее, СТГ оказывает определенное модулирующее влияние и на активность аденилатциклазы.
Другие гипофизарные гормоны (АКТГ, ТТГ, меланоцитостимулирующий гормон) также оказывают липолитическое действие, хотя и менее выраженное, чем у СТГ.
Тиреоидные гормоны и кортикостероиды оказывают пермиссивное (разрешающее) действие на липолиз, поскольку липолитический и калоригенный эффекты катехоламинов не проявляются в отсутствие кортикостероидов и тиреоидных гормонов. Имеются данные о том, что ТТГ, АКТГ, гормоны щитовидной железы и коры надпочечников индуцируют синтез аденилатциклазы и, следовательно, принимают непосредственное участие в стимуляции липолиза.
Как уже отмечалось, инсулин обладает характерным антилиполитическим свойством, и при сахарном диабете вследствие увеличения липолиза повышается концентрация свободных жирных кислот в плазме, снижению которой способствует инсулинотерапия.
Белковый обмен.
Белки в организме также являются источником энергии. Они содержатся главным образом в мышцах и их количество составляет в организме здорового человека массой 70 кг около 6000 г, что соответствует 24 000 ккал. Циркуляция их в крови в виде аминокислот незначительна и составляет всего 6 г, или 24 ккал.
По воздействию на белковый обмен все гормоны можно разделить на две большие группы: 1) гормоны, первичное действие которых – стимуляция процессов транскрипции – проявляется в ядре клетки (стероидные и тироидные гормоны); 2) гормоны, стимулирующие процессы трансляции, происходящие в цитоплазме клетки.
Анаболическое действие инсулина заключается в ускорении проникновения аминокислот через мембрану клетки и включения их в белки, что вызывает снижение уровня аминокислот в крови. Это действие инсулина не зависит от синтеза РНК. и не угнетается актиномицином. Инсулин снижает активность аминотрансфераз и ферментов цикла мочевины. Последний эффект инсулина характеризуется повышением активности РНК-полимеразы и концентрации РНК в печени. При этом увеличивается скорость образования полисом и рибосом.
Действие инсулина на синтез белка в печени противоположно действию глюкагона, который усиливает катаболизм аминокислот и мобилизует их для процессов глюконеогенеза.
Основным эффектом действия гормона роста является стимуляция анаболичесих процессов, чем и обусловливается ускорение роста скелета при гиперсекреции этого гормона. Он, так же как инсулин, стимулирует транспорт аминокислот и их включение рибосомами в белки, повышает скорость образования полисом и рибосом, активность РНК-полимеразы, увеличивает количество РНК в клетках, что сопровождается повышением скорости синтеза белков. Гормон роста угнетает активность ферментов, разрушающих аминокислоты, повышает уровень инсулиноподобных факторов роста I и II (ИФР I и II) в сыворотке крови, стимулирует синтез коллагена в костях, коже, других органах и тканях организма.
Тиреоидные гормоны не влияют на транспорт аминокислот. Под действием этих гормонов возрастает активность ферментов, в том числе глицеролфосфатдегидрогеназы, цитохромоксидазы, цитохрома С, NAD-изолимонной дегидрогеназы, а также РНК-полимеразы и увеличивается синтез РНК (включая м-РНК).
Под влиянием эстрогенов увеличивается содержание РНК, что приводит к усилению синтеза белка.
Андрогены оказывают на белковый обмен такое же действие, как и эстрогены. Кроме того, тестостерон и другие андрогены влияют на развитие мышечной ткани, улучшая транспорт аминокислот и повышая активность РНК-полимеразы.
Таким образом, гормоны регулируют все виды обмена веществ (углеводный, жировой, белковый, минеральный и др.), поддерживая постоянство внутренней среды, которое, как установил еще Клод Бернар, является непременным условием жизнедеятельности организма.
Дата добавления: 2018-11-25; просмотров: 968;