Сегментарная реакция
развивается по принципу сомато-висцерального рефлекса, возникающего в первую очередь в зоне соответствующего кожного метамера. Известно, что большинство афферентов от внутренних органов поступают в ЦНС на нескольких уровнях — полисегментарно и организованы по типу двусторонней воронки: в одном органе перекрываются афференты из различных сегментов, а в одном сегменте перекрываются пути, идущие от разных внутренних органов. При этом включаются звенья рефлекторной дуги спинного мозга, ствола головного мозга (при воздействии на ТА лица и головы), сегментарных образований вегетативной нервной системы.
Эфферентная часть реакции осуществляется в виде возбуждения двигательных и вегетативных нервных проводников с ответом со стороны внутренних органов, поперечнополосатых мышц и кожи (висцеро-мышечно-кожные рефлексы).
Степень выраженности сегментарной реакции зависит от интенсивности стимула, его характеристики (возбуждающий или тормозной метод воздействия), от исходного состояния внутреннего органа и сегментарной специфичности ТА. Показано [95], что стимуляция поверхности брюшной стенки, вентральной и дорсальной поверхностей грудной клетки приводит к выраженному угнетению перистальтики желудка. Аналогичная стимуляция нижней челюсти или ноги её усиливает. Афферентное звено этих рефлексов состоит из соматических волокон соответствующих частей тела, а эфферентным звеном в одних случаях является симпатический нерв желудка, в других — ветвь блуждающего нерва. Соответственно переключающая часть рефлекторных дуг может располагаться как на сегментарном, так и супрасегментарном уровне.
Сегментарная реакция зависит не только от места приложения раздражения, но и от исходного состояния внутреннего органа. Так, стимуляция промежности при пустом или слегка растянутом мочевом пузыре вызывает его сокращение. При растянутом мочевом пузыре такая же стимуляция вызывает угнетение его естественных сокращений. Аналогичные закономерности в отношении места раздражения и состояния внутреннего органа выявлены при исследованиях влияния соматической стимуляции на мозговой кровоток сомато-сердечного, сомато-адреналового и сомато-селезёночных рефлексов.
Важное значение имеет и характер наносимого раздражения. Например, неболевая тактильная стимуляция в области груди или задних конечностей вызывает снижение импульсации в надпочечниковом эфферентном симпатическом нерве и уменьшение секреции катехоламинов мозговым веществом надпочечников. Ноцицептивная стимуляция этих же зон приводит к противоположным эффектам. Результаты приведённых экспериментальных исследований подтверждают эмпирически установленное важное значение основных факторов РТ — Места, Момента и Метода раздражения.
Большинство передаточных нейронов дорсальных рогов спинного мозга получают входы как от соматических, так и висцеральных нервных волокон, что обеспечивает простейший механизм взаимного подавления сигналов, приходящих по этим волокнам. Нейрон, занятый ответом на сигнал, поступивший по кожному волокну, просто не имеет возможности ответить на сигнал, пришедший от внутреннего органа.
Болевые (ноцицептивные) сигналы, поступающие по тонким миелинизированным (AS) и немиелинизированным (С) волокнам, конкурируют с неболевыми сигналами, поступающими по толстым миелиновым волокнам (А? и Ау) за возможность возбудить их общий передаточный нейрон (Т). Схема их конкурирующих взаимоотношений с участием нейронов желатинозной субстанции (ЖС) была предложена Melzack и Woll [131] и получила название теории воротного контроля боли (рис.4).
Обозначения:
Ab, Ay— толстые миелинизированные афферентные волокна AS—тонкие миелинизированнные ноцицептивные волокна С — тонкие немиелинизированные ноцицептивные волокна ЖС — нейрон желатинозной субстанции Т — передаточный (трансмиттерный) нейрон
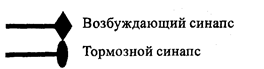
Рис. 4. Схема воротного контроля боли Melzack и Woll
42
Сигналы, поступающие по А8 и С-волокнам, тормозят вставочный нейрон ЖС, в результате чего снимается его пресинаптическое торможение передаточного нейрона Т — «ворота открываются». Ар и Ау волокна возбуждают нейрон ЖС, в результате чего усиливается его пресинаптическое торможение передаточного нейрона Т — «ворота закрываются». В настоящее время установлено, что торможение А8 и С-волокнами нейронов ЖС осуществляется с участием субстанции П, а в возбуждении его А? и Ау волокнами участвуют энкефалины и динорфин. Таким образом, регуляторные пептиды, участвующие в периферических механизмах развития гипералгезии и антиноцицепции, воспроизводят своё действие и на уровне ЦНС.
Механизм воротного контроля боли удовлетворительно объясняет обезболивающее действие стимулов, активирующих толстые миелиновые волокна. Учитывая морфологические данные о высокой плотности в ТА таких волокон и рецепторов, с ними связанных, а также возбуждение их в результате локального мышечного спазма под влиянием введённой иглы, развитие рефлекторной аналгезии (РА) объясняют механизмом воротного контроля. Однако, при стимуляции ТА возбуждаются не столько толстые миелиновые волокна, сколько тонкие миелиновые и безмиелиновые волокна, проводящие эпикритические и протопатические болевые ощущения, поэтому теория воротного контроля не может служить вполне адекватным объяснением общего механизма акупунктурной аналгезии.
Нейрофизиологические механизмы сегментарного уровня лежат в основе реципрокного принципа лечения, используемого в традиционной АЛ — «верх лечи через низ, а низ — через верх; левую сторону лечи через правую, а правую — через левую».
Принципу реципрокности отвечает правило «полдень-полночь», согласно которому воздействуют на точки АК, находящегося по отношению к поражённому в противоположной фазе суточной активности. Правило «полдень — полночь» связывает АК верхних и нижних конечностей. Воздействие на ТА возбуждающим или тормозным методом приводит к одноимённому изменению состояния возбудимости мотонейронов на стороне воздействия и к противоположным изменениям их возбудимости на контрлатеральной стороне.
Такая же инверсия направленности реакции происходит при воздействии на ТА, находящиеся в мышцах-антагонистах. Например, укалывание точки BL(V) 60 (кунь-лунь), расположенной у латеральной лодыжки, вызывало снижение возбудимости мотонейронов медиальной головки икроножной мышцы, а такая же стимуляция точки KI(R) 5 (тай-си), расположенной у медиальной лодыжки, — повышение их возбудимости. Выявлены 4 варианта реципрокности, соответствующих генератору локомоторной активности. При первом варианте изменения возбудимости мотонейронов и рецепторов происходят в мышцах-антагонистах всех 4-х конечностей в противофазном режиме.
Второй вариант реципрокных координации (феномен иноходца) отличается одинаковой реакцией мотонейронов мышц-сгибателей или разгибателей и рецепторов, покрывающих их кожную поверхность в конечностях одной половины тела. Третий вариант — рострально-каудальная, или верхне-нижняя реципрокность (феномен галопа). Четвёртый вариант — дорсо-вентральной (передне-задней) реципрокности состоит в однонаправленном изменении возбудимости мотонейронов и рецепторов над всей разгибательной (янской) поверхностью и в противоположном направлении над сгибательной (иньской) поверхностью тела. Исходя из указанных вариантов сопряжённых реципрокных координации, выбирают место, момент и метод акупунктурного воздействия [ИЗ].
Общая реакция
развивается вследствие поступления афферентых сигналов от периферических рецепторов по спинномозговым соматическим и вегетативным путям в ретикулярную формацию ствола, подкорковые и корковые отделы ЦНС с изменением их функционального состояния и развитием комплекса нейрогуморальных, вегетативных и эмоциональных адаптационных реакций.
Функциональные взаимоотношения между кожными покровами и внутренними органами складывались в процессе эволюции. Ведущую роль при этом играли рецепторы наружных оболочек, подвергающиеся непосредственным воздействиям факторов внешней среды, особенно те из них, которые сигнализировали о возможном повреждении (ноцицепторы). Необходимость обеспечения сначала простейших, а затем и более сложных адаптационных реакций, требовала формирования регуляторных связей между экстерорецепторами и внутренними органами.
Поскольку никакая регуляция невозможна без обратной связи, в филогенезе формировались вначале двусторонние функциональные взаимосвязи между внутренними органами и кожными покровами по метамерному принципу. Затем в связи с развитием головного мозга и дальнейшим процессом дифференцировки периферической нервной системы эти связи становились более сложными.
Онтогенез повторяет основные стадии филогенеза. Структурно-функциональные особенности сомато-висцеральных и висцеро-соматических механизмов регуляции закладываются на ранних этапах эмбриогенеза. Кожа и нервная система формируются из эктодермы, а внутренние органы — из мезо- и эндодермы. Процессы дифференцировки зародышевых листков обеспечивают взаимосвязь экто-дермального нейротома с мезодермальными склеротомом, миотомом и дерматомом и мезо-энтодермальным спланхнотомом.
Изначально организуемые по метамерному принципу, эти взаимосвязи видоизменяются и отчасти зашифровываются в результате неравномерного развития и перемещения органов в процессе превращения зародыша в плод. При этом, однако, сохраняется их исходное сродство, что проявляется в дистантных, внесегментарных взаимосвязях ТА с определёнными внутренними органами. Указанные положения составляют основу «эмбриональной» теории АП, предложенную R. de la Fuye в 1956 г. и получившую дальнейшее развитие [5].
Исходное близкое соседство зачатков органов и тканей в раннем эмбриогенезе обеспечивает их биохимическую родственность в постнатальном периоде и во всём дальнейшем онтогенезе. Под биохимической родственностью подразумевается прежде всего наличие на клетках единых типов и классов молекулярных рецепторов, которые обеспечивают существование регуляторного континуума. Очевидно, клетки взрослого организма, располагающиеся в различных сегментах тела, но происходящие из предшественников, соседствующих друг с другом в эмбриональном периоде, имеют больше шансов иметь одинаковый набор мембранных рецепторов, обеспечивающих их более тесное, прежде всего информационное взаимодействие как с участием, так и без непосредственного участия нервной системы.
С помощью регуляторного континуума может осуществляться отражение патологического процесса, происходящего в части тела или внутреннем органе, метамерно не связанных с репрезентативной AT. Например, содержание регуляторных пептидов в аурикулярных точках, соответствующих поясничному отделу позвоночника, коррелирует с выраженностью клинических проявлений пояснично-крестцового радикулита.
Дата добавления: 2017-12-05; просмотров: 1102;