ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА, СТРОЕНИЕ И ФУНКЦИИ. СТРУКТУРЫ, ФОРМИРУЕМЫЕ ПЛАЗМАТИЧЕСКОЙ МЕМБРАНОЙ
Мы начнем гистологию с изучения эукариотической клетки, являющейся самой простой системой, наделенной жизнью. При исследовании клетки в световом микроскопе мы получаем информацию о ее размере, форме, и эта информация связана с наличием у клеток ограниченных мембраной границ. С развитием электронной микроскопии (ЭМ) наши представления о мембране, как о четко ограниченной линии раздела между клеткой и окружающей средой изменились, ибо оказалось, что на поверхности клетки имеется сложная структура, состоящая из следующих 3-х компонентов:
1. Надмембранный компонент (гликокаликс) (5 - 100 нм);
2. Плазматическая мембрана (8 - 10 нм);
3. Подмембранный компонент (20 – 40 нм).
При этом 1 и 3 компоненты вариабельны и зависят от типа клеток, наиболее статичным представляется строение плазматической мембраны, которую мы и рассмотрим.
Плазматическая мембрана. Изучение плазмолеммы в условиях ЭМ привело к заключению об однотипности ее структурной организации, при которой она имеет вид триламинарной линии, где внутренний и наружный слои электронноплотные, а расположенный между ними – более широкий слой представляется электроннопрозрачным. Такой тип структурной организации мембраны свидетельствует об ее химической гетерогенности. Не касаясь дискуссии по этому вопросу, оговорим, что плазмолемма состоит из трех типов веществ: липидов, белков и углеводов.
Липиды, входящие в состав мембран, обладают амфифильными свойствами за счет присутствия в их составе как гидрофильных, так и гидрофобных групп. Амфипатический характер липидов мембраны способствует образованию липидного бислоя. При этом в фосфолипидах мембраны выделяют два домена:
а) фосфатная– голова молекулы, химические свойства этого домена определяют его растворимость в воде и его называют гидрофильным;
б) ацильные цепи, представляющие собой этерифицированные жирные кислоты – это гидрофобный домен.
Типы мембранных липидов: основным классом липидов биологических мембран являются фосфолипиды, они формируют каркас биологической мембраны. См. рис.1
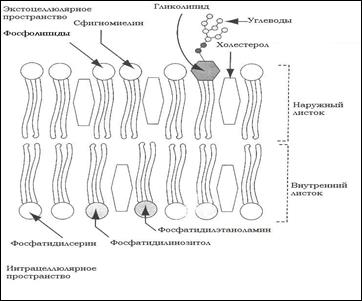
Рис. 1: Типы мембранных липидов
Биомембраны – это двойной слой амфифильных липидов(липидный бислой). В водной среде такие амфифильные молекулы самопроизвольно образуют бислой, в котором гидрофобные части молекул ориентированы друг к другу, а гидрофильные к воде. См. рис. 2
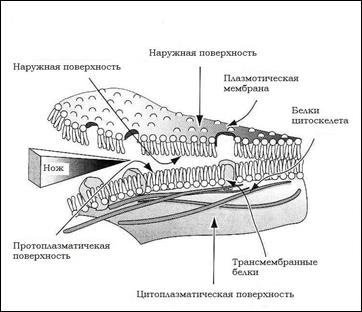
Рис. 2: Схема строения биомембраны
В состав мембран входят липиды следующих типов:
1. Фосфолипиды;
2. Сфинголипиды - “головки” + 2 гидрофобных “хвоста”;
3. Гликолипиды.
Холестерин (ХЛ)– находится в мембране в основном в срединной зоне бислоя, он амфифилен и гидрофобный (за исключением одной гидроксигруппы). Липидный состав влияет на свойства мембран: отношение белок/липиды близок 1:1, однако миелиновые оболочки обогащены липидами, а внутренние мембраны – белками.
Способы упаковки амфифильных липидов:
1. Бислои (липидная мембрана);
2. Липосомы - это пузырек с двумя слоями липидов, при этом как внутренняя, так и наружная поверхности являются полярны;
3. Мицеллы – третий вариант организации амфифильных липидов – пузырек, стенка которого образована одним слоем липидов, при этом их гидрофобные концы обращены к центру мицеллы и их внутренняя среда является не водной, агидрофобной.
Наиболее распространенной формой упаковки молекул липидов является образование ими плоского бислоя мембран. Липосомы и мицеллы – это скорые транспортные формы, обеспечивающие перенос веществ в клетку и из нее. В медицине липосомы используют для переноса водорастворимых, а мицеллы – для переноса жирорастворимых веществ.
Белки мембраны
1. Интегральные (включены в липидные слои);
2. Периферические. См. рис. 3
Интегральные (трансмембранные белки):
1. Монотопные– (например, гликофорин. Они пересекают мембрану 1 раз), и являются рецепторами, при этом их наружный – внеклеточный домен – относится к распознающей части молекулы;
2.Политопные– многократно пронизывают мембрану – это тоже рецепторные белки, но они активизируют путь передачи сигнала внутрь клетки;
3.Мембранные белки, связанные с липидами;
4. Мембранные белки, связанные с углеводами.
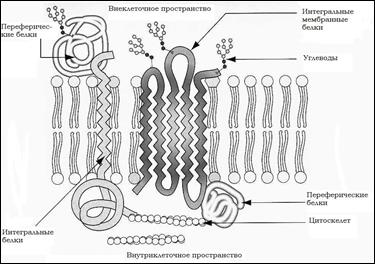
Рис. 3: Белки мембраны
Периферические белки:
Не погружены в липидный бислой и не соединены с ним ковалентно. Они удерживаются за счет ионных взаимодействий. Периферические белки ассоциированы с интегральными белками в мембране за счет взаимодействия - белок-белковые взаимодействия.
1. Спектрин, который расположен на внутренней поверхности клетки;
2.Фибронектин, локализован на наружной поверхности мембраны.
Белки – обычно составляют до 50% массы мембраны. При этом интегральные белкивыполняют следующие функции:
а) белки ионных каналов;
б) рецепторные белки.
А периферические мембранные белки (фибриллярные, глобулярные) выполняют следующие функции:
а) наружные (рецепторные и адгезионные белки);
б) внутренние – белки цитоскелета (спектрин, анкирин), белки системы вторых посредников.
Ионные каналы– это сформированные интегральными белками каналы, они формируют небольшую пору, через которую по электрохимическому градиенту проходят ионы. Наиболее известные каналы – это каналы для Nа, К, Са, Сl.
Существуют и водные каналы – это аквопорины (эритроциты, почка, глаз).
Надмембранный компонент – гликокаликс, толщина 50 нм. Это углеводные участки гликопротеинов и гликолипидов, обеспечивающие отрицательный заряд. Под ЭМ – это рыхлый слой умеренной плотности, покрывающий наружную поверхность плазмолеммы. В состав гликокаликса помимо углеводных компонентов входят периферические мембранные белки (полуинтегральные). Функциональные участки их находятся в надмембранной зоне - это иммуноглобулины. См. рис. 4
Функция гликокаликса:
1. Играют рольрецепторов;
2. Межклеточное узнавание;
3. Межклеточные взаимодействия (адгезивные взаимодействия);
4. Рецепторы гистосовместимости;
5. Зона адсорбции ферментов (пристеночное пищеварение);
6.Рецепторы гормонов.
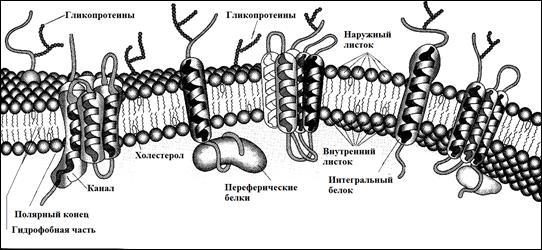
Рис. 4: Гликокаликс и подмембранные белки
Подмембранный компонент - самая наружная зона цитоплазмы, обычно обладает относительной жесткостью и эта зона особенно богата филаментами (d = 5-10 нм). Предполагают, что интегральные белки, входящие в состав клеточной мембраны, прямо или косвенно связаны с актиновыми филаментами, лежащими в подмембранной зоне. При этом экспериментально доказано, что при агрегации интегральных белков, находящийся в этой зоне актин и миозин также агрегируют, что указывает на участие актиновых филамент в регуляции формы клетки.
| <== предыдущая лекция | | | следующая лекция ==> |
| РЫХЛАЯ ВОЛОКНИСТАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ | | |
Дата добавления: 2017-09-19; просмотров: 3673;