Эволюция протобионтов
Первичные организмы на Земле были гетеротрофами, так как использовали в качестве пищи органические вещества первичного бульона. Они были способны утилизировать самые различные восстановленные субстраты, которых должно было быть немало 4 млрд лет назад на поверхности Земли (Н2, H2S, F++, NH3, NO-2, абиогенная органика). Уже на самых ранних этапах эволюции органического мира должен был возникнуть процесс фотофосфорилирования (В.П. Скулачев). При этом первичные фотосинтетики не производили кислорода. Этот этап эволюции сегодня представлен зелеными и пурпурными бактериями.
2,5 млрд лет назад с возникновением цианобактерий положение изменилось. В результате фотосинтеза впервые стал возникать и накапливаться в атмосфере Земли кислород (рис.2).
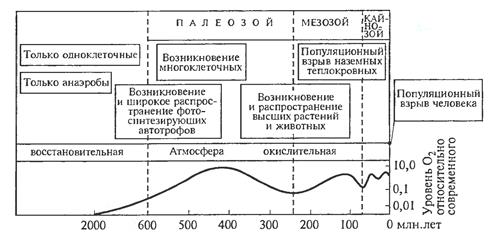
Рисунок 2- Эволюция биосферы и атмосферы( Ю. Одум, 1975)
Благодаря фотосинтезу в каждый последующий этап в органическом веществе, находящемся на поверхности Земли, накапливалось все больше и больше энергии солнечного света. Это способствовало ускорению биологического круговорота веществ и ускорению эволюции в целом.
Около 2,5 – 2,0 млрд лет назад концентрация кислорода в атмосфере достигла 1 % современной. Это сделало возможным появление более крупных, чем бактериальные, эукариотных организмов (Б.М. Медников).
Большинство исследователей вслед за Карлом Вёзе признают, что вскоре после возникновения жизни она разделилась на тринадцарства (домена). По-видимому, больше всего черт исходных протоорганизмов сохранили архебактерии. Они обитают в бескислородных илах, концентрированных растворах солей, горячих вулканических источниках. Второе мощное надцарство — эубактерии. Из третьего корня развилась ветвь организмов, имеющих оформленное ядро с оболочкой,— эукариоты.
По устаревшей классификации, имеются два надцарства (империи): прокариоты (включая царства архебактерий и эубактерий) и эукариоты. Иногда к ним добавляли ещё одно надцарство акариот (вирусов).
Есть обоснованная гипотеза (эндосимбиоза), что эукариоты возникли в результате симбиоза их предков с предками митохондрий и хлоропластов — аэробных бактерий и цианобактерий (рис. 3). Эта гипотеза дает удовлетворительное объяснение многим чертам сходства в строении и биохимических особенностях органелл — внутриклеточных источников энергии эукариот,— с таковыми свободноживущих прокариот.
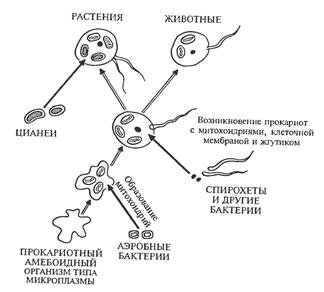
Рисунок 3 – Схема этапов симбиогенеза при возникновении основных групп живых
организмов (А.Л.Тахтаджян, 1972).
Колоссальное значение для развития биосферы в целом имело возникновение и распространение одной из групп эубактерий — цианобактерий. Они способны осуществлять оксигенный фотосинтез, и в результате их жизнедеятельности в атмосфере Земли должен был появиться кислород в достаточно больших количествах. Появление кислорода в атмосфере определило возможность последующего развития растений и животных.
Надцарство эукариот очень рано, более чем миллиард лет назад, разделилось на царства животных, растений и грибов. Грибы более близки к животным, чем к растениям. Наконец, небольшая группа слизевиков настолько своеобразна, что его выделяют в отдельное царство в надцарстве эукариот. По-видимому, многоклеточность возникла независимо у грибов, растений, кишечнополостных и других животных.
4. Основные этапы эволюции биосферы в целом
Первичные экосистемы были, несомненно, связаны с водой. В последние годы внимание микробиологов привлекли цианобактериальные «маты» — пласты микробного происхождения, располагающиеся на ровных увлажненных пространствах в некоторых прибрежных районах. Предполагается, что подобные маты могли представлять первый живой покров нашей планеты в докембрии и явились основой для развития почвы в будущем. Возможно, тогда существовал единый цианобактериальный мат «от моря и до моря» (Г.А.Заварзин) — первая наземная экосистема планеты.
В процессе эволюции биосферы очень скоро определилась ее «двухслойная» структура — бактериально - микробное «основание» и эукариотная «надстройка» (Г.А. Заварзин). «Основание» неизмеримо более устойчиво, и даже в наше время спустя 3,9 млрд лет после начала эволюции биосферы мы обнаруживаем точно такие же микробные сообщества, которые были характерны для ранних этапов эволюции биосферы (в основном вокруг горячих источников — гидротермов — на суше и в глубинах океана). Это бактериально - микробное основание биосферы было преобладающим на протяжении сотен миллионов лет (табл.1).
Первые растения — зеленые водоросли — обнаружены в отложениях архея (около 3 млрд лет назад). Первые животные пока в ископаемом виде не найдены, но в верхнем протерозое, особенно в венде (около 1,5 млрд лет назад), достоверно обнаруживаются многоклеточные кишечнополостные, членистоногие и иглокожие. В венде же появились первые растительноядные организмы (амебы, инфузории, грибы), что сразу же усложнило существовавшие прежде экосистемы консументами первого порядка.
Возникновение многоклеточных организмов сопровождалось повышением устойчивости экосистем и открыло возможности для их развития в разных направлениях.
В начале палеозоя, в кембрии, сразу в нескольких группах организмов возникают минерализованные скелеты (из кремния, стронция, кальция, апатита, хитина, спонгина). Эта «скелетная революция» (Б.М. Медников) могла означать только появление в экосистемах консументов второго порядка — хищников, что было одной из главных причин резкого увеличения многообразия экосистем. Другая возможная причина — появление более совершенных организмов - фильтраторов. Без существования таких живых фильтров водоемы, включая Мировой океан, были бы очень мутными, с совершенно иными физико-химическими характеристиками. Органическое вещество, транспортируемое в фекалиях (пеллетах) на дно водоемов, в результате работы фильтраторов обогащало донные осадки, создавало условия для развития богатых донных экосистем. Это было причиной развития многообразия трилобитов и приапулид. Только начиная с ордовика — следующего за кембрием периода палеозоя (табл. 1)—начинает формироваться разнообразная фауна экосистем, связанная с освоением самой толщи воды.
С освоением водоемов живыми организмами гидросфера планеты превращается из косной в биокосную.
Появление и распространение сосудистых растений в силуре и девоне сыграло важную роль не только для дальнейшего изменения газового состава атмосферы, но и для уменьшения эрозии суши и стабилизации линии раздела суши и водоемов. Возникновение наземных растений, способных к вертикальному росту, позволило фотосинтезирующим структурам биосферы располагаться в трехмерном пространстве, что резко интенсифицировало весь процесс фотосинтеза. Развитие наземной растительности привело к существенному усложнению наземных экосистем (развитие почвы, корневой системы, гниющей растительной подстилки, накопление больших запасов биомассы).
Но в то же время потребовались сотни миллионов лет, чтобы в экосистемах того периода возникло достаточное число консументов зеленой массы. Поэтому огромное количество органического вещества оказывалось непотребленным и выводилось из биотического круговорота в виде дошедших до нас биогенных горючих ископаемых — каменного угля, нефти, горючих сланцев.
Таблица 1- Геохронологическая и стратиграфическая шкала Земли
| Эон | Эра | Период (система) | Эпоха (отдел) | Изотопные датировки, млн лет | Характерные формы жизни |
| Фанерозой | Кайнозой | Четвертичный | Голоцен Плейстоцен | 1.8 | Развитие рода Homo |
| Неоген | Плиоцен | 25 ±2 | Возникновение современных семейств млекопитающих, современной флоры | ||
| Палеоген | Миоцен Олигоцен Эоцен Палеоцен | 66 ±3 | Расцвет млекопитающих и птиц | ||
| Мезозой | Мел | 136 + 5 | Развитие цветковых растений, расцвет насекомых, вымирание многих рептилий | ||
| Юра | 190-195±5 | Господство рептилий на суше, в воде и воздухе. Возникновение покрытосеменных и птиц | |||
| Триас | 230 ±10 | Расцвет рептилий, распространение голосеменных, появление млекопитающих | |||
| Палеозой | Пермь | Появление голосеменных, распространение рептилий | |||
| Карбон | 280 ±10 | Распространение лесов, расцвет амфибий, возникновение летающих насекомых и рептилий | |||
| Девон | 345 ±10 | Господство рыб, возникновение насекомых и амфибий, появление лесов из папоротников и плаунов | |||
| Силур | 400 ±10 | Выход растений и беспозвовоночных на сушу | |||
| Ордовик | 435 ±10 | Появление первых позвоночных — бесчелюстных | |||
| Кембрий | 490 +15 | Развитие беспозвоночных, появление высших растений | |||
| Криптозой | Венд | 570 ±20 | Появление кишечнополостных, членистоногих, иглокожих | ||
| Протерозой | Рифей | 650-690±20 | Появление эукариот, многоклеточных растений и животных | ||
| Карелий | 1650 ±50 | Развитие низших растений | |||
| Архей | 2500 ±100 | Господство бактерий и синезеленых, появление зеленых водорослей | |||
| 3500 - 3900 | Зарождение жизни, появление прокариот. |
Одним из революционных моментов в дальнейшем развитии экосистем биосферы Земли явилось, по-видимому, возникновение в середине мелового периода однолетних травянистых растений. Они в сочетании с быстро эволюционировавшими вслед за ними животными и грибами способствовали резкому ускорению круговорота биогенов в экосистемах.
На границе мезозоя и палеозоя произошло резкое сокращение биоразнообразия морской биоты — число семейств морских организмов уменьшилось за несколько десятков миллионов лет вдвое. Причины этого пока неясны, тем более что такого кризиса в это время не прослеживается в наземных экосистемах.
Впоследствии, в мезозое и кайнозое, неоднократно наблюдались кризисы биоразнообразия меньшего масштаба. Тогда на протяжении исторически коротких отрезков времени (нескольких миллионов лет) исчезло до четверти ранее встречавшихся таксонов. Есть немало предположений о причинах таких кризисов, но ни одно из них не может считаться достаточно доказанным и исчерпывающим.
В процессе эволюции жизни на Земле важным шагом было образование биотического круговорота — создание таких экосистем, в которых одна и та же порция вещества могла многократно использоваться. Это стало возможным, когда экосистемы включили не только организмы-автотрофы (хемо- и фотосинтетиков), но и гетеротрофы. Так возникла вся триада продуцентов (производителей) — консументов (потребителей) — редуцентов (разрушителей, минерализаторов). Важнейшим направлением дальнейшей эволюции экосистем стало уменьшение потери вещества из биотических круговоротов, а также интенсификация миграции химических элементов.
Таким образом, в процессе развития жизни на нашей планете в целом происходило усложнение экосистем, сопровождающееся возрастанием числа одновременно существующих видов, экспансией жизни, охватывающей ныне всю поверхность планеты, усиливающейся дифференциацией этой живой оболочки на биогеографические области, биомы, формации. Результатом миллиардов лет эволюции экосистем является современная биосфера Земли, дифференцированная на множество биогеоценозов и включающая более двух миллионов ныне живущих видов. Одним из результатов дифференциации биосферы в ходе эволюции должно было быть и повышение устойчивости биосферыв целом.
Итак, основными чертами эволюции биосферы Земли в целом были следующие:
—общее усложнение экосистем и возрастание «суммы жизни»;
—возникновение биотического круговорота веществ, осуществляемого с помощью цепочки взаимосвязанных продуцентов — консументов — редуцентов;
—накопление массы органического вещества;
—все более полное освоение живым веществом косного, возникновение биокосных экосистем (почва, современный океан);
—циклический характер нарастания биоразнообразия.
5. Основные пути эволюции растений
Число видов ныне существующих растений достигает более 500 тыс., из них цветковых примерно 300 тыс. видов. Остатки зеленых водорослей находят в породах архейского возраста (около 3 млрд лет назад). В протерозое в морях обитало много разных представителей зеленых и золотистых водорослей. В это же время, видимо, появились первые прикрепленные ко дну водоросли. Первичные почвообразовательные процессы в протерозое подготовили условия для выхода настоящих растений на сушу. В первые периоды палеозоя растения населяют в основном моря. Среди прикрепленных ко дну встречаются зеленые и бурые водоросли, а в толще воды — диатомовые, золотистые, красные и другие водоросли.
В силуре (435—400 млн лет назад) в царстве растений происходит крупное эволюционное событие: растения (риниофиты) выходят на сушу.
Первичные фототрофные низшие растения были также разнообразны по составу, среди них были группы с наличием хлорофилла «а» и «b», с высоким содержанием каротиноидов и хлорофилла «с» и, наконец, группы с преобладанием фикобилинов. Вероятно, между этими группами организмов не было генетического единства. Разнообразие состава первичных фототрофов было вызвано, несомненно, достаточно разнородными условиями существования и позволяло полнее использовать особенности среды.
В конце силура отмечено появление первых по-настоящему наземных растений — псилофитов, которые покрывали сплошным зеленым ковром прибрежные участки суши. Это было важным эволюционным шагом. Происходит перестройка в проводящей системе и покровных тканях: у псилофитов появляются проводящая сосудистая система со слабо дифференцированной ксилемой и флоэмой, кутикула и устьица. Псилофиты оказались и более надежно прикрепленными к субстрату с помощью дихотомически разветвленных нижних осей: у некоторых из них обнаружены примитивные «листья». Псилофиты занимали промежуточное положение между наземными сосудистыми растениями и водорослями: внешне были похожи на водоросли, тело не было дифференцировано на вегетативные органы и имело большую испаряющую поверхность.
Дальнейшая эволюция растений в наземных условиях привела к усилению компактности тела, появлению корней, развитию эпидермальной ткани с толстостенными, пропитанными восковидным веществом клетками, замене трахеид сосудами, изменению способов размножения, распространения и т. д. Переход от трахеид к сосудам — приспособление к засушливым условиям; с помощью сосудов создается возможность быстрого поднятия воды на большие высоты. Переход к сосудам начался в корнях, стеблях, затем в листьях.
Начальные этапы эволюции наземных растений связаны с возникновением архегониальных форм — мохообразных, папоротникообразных и голосеменных. У всех этих групп женский половой орган представлен архегонием, а мужской — антеридием. Предполагают, что архегониальные произошли от бурых или зеленых водорослей. При выходе на сушу защита гаметангиев водорослей от иссушения обеспечивалась преобразованием их в архегоний и антеридий. Этому способствовали изменение формы гаметангиев и образование у них многослойных стенок.
С момента выхода на сушу растения развиваются в двух основных направлениях: гаметофитном и спорофитном. Гаметофитное направление было представлено мохообразными, а спорофитное — остальными высшими растениями, включая цветковые. Спорофитная ветвь оказалась более приспособленной к наземным условиям: особого развития достигает корневая система, усложняется и совершенствуется проводящая система, заметно совершенствуются покровные и механические ткани, а также способы размножения и создаются возможности для снижения частоты проявления возникающих летальных и других мутаций (в результате диплоидизации организма). В наземных условиях оказались непригодными для размножения свободно плавающие незащищенные половые клетки, здесь для целей размножения формируются споры (семена), разносимые ветром.
Уже в девоне встречаются пышно развитые леса из прогимноспермов, папоротников и плаунов. Эти леса еще более распространяются в карбоне, характеризующемся увлажненным и равномерно теплым климатом в течение всего года. Мощные споровые растения — лепидодендроны и сигиллярии — достигали 40 м высоты. В карбоне же получили развитие первые семенные — голосеменные: птеридоспермы, древесные кордаиты и гинкговые, часть из которых вымирает в перми, около 280 млн лет назад.
Генеральная линия эволюции папоротникообразных на суше шла по пути преобразования спорофита (бесполое поколение). Он достиг совершенства как по разнообразию форм (деревья и травы), так и по строению. В засушливых условиях гаметофит (половое поколение) стал уже помехой из-за необходимости капельно-жидкой воды для переноса гамет. Поэтому не удивительны редукция гаметофита и значительное развитие спорофита в ходе дальнейшей эволюции наземных растений (рис.4).
Одним из важных эволюционных приобретений считается появление разноспоровых папоротников, предшественников семенных растений. Начиная с лепидодендрона у некоторых плауновидных (селагинелла), папоротников и семенных папоротников закрепляется разноспоровость; в пазухах спорофитов развиваются мега- и микроспорангии. Такое событие отмечено в силуре — девоне, т. е. примерно 400 млн лет назад. Мегаспорангии имели 4 мегаспоры, а микроспорангии — множество микроспор. Дифференциация спорангиев и спор привела к появлению разных размеров гаметофитов (включая и очень мелких) и разобщению мужского и женского гаметофитов, что в конечном итоге оказало влияние на редукцию гаметофита (гаплоидного тела). Редукция гаметофита способствовала удлинению диплоидной фазы развития организма, удлинению и усложнению процессов дифференциации и онтогенеза. Не случайно, что первые разноспоровые достигали гигантских размеров; это сигиллярии, лепидодендроны, гигантские папоротники, каламиты.
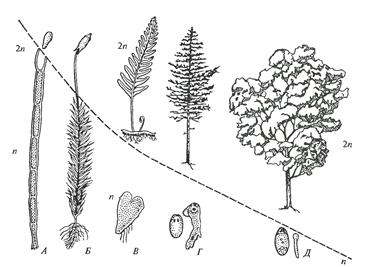
Рисунок 4 – Схема эволюционных изменений растений в направлении увеличения размеров и значения бесполого поколения (2n), а также редукции размеров полового поколения (n): А-водоросли, Б-мхи, В-папоротники, Г-голосеменные, Д- покрытосеменные (К. Фуллер, О.Типпе, 1954)
Важнейшее событие в жизни голосеменных — превращение мегаспорангия в семязачаток, семяпочку с защитными покровами — интегументами и полное освобождение у всех семенных процесса полового размножения от воды. Микроспорангии у голосеменных превращаются в гнезда пыльника. Спермии большинства голосеменных неподвижны, и перенос их к архегониям осуществляется пыльцевой трубкой. Потеря мужским гаметофитом самостоятельности привела к редукции его до пыльцевой трубки с вегетативным ядром и двумя сперматозоидами. Опыление у голосеменных осуществляется ветром и нередко насекомыми, после оплодотворения семязачаток превращается в семя. Заметим, что семя появляется у семенных папоротников еще в девоне, т. е. задолго до развития цветка. Переход к семенному размножению связан с рядом эволюционных преимуществ; диплоидный зародыш в семенах защищен от неблагоприятных условий наличием покровов и обеспечен пищей, а семена имеют приспособления для распространения животными и др. Эти и другие преимущества способствовали широкому распространению семенных растений.
Непосредственные предки покрытосеменных пока не найдены в ископаемом виде. Считается, что покрытосеменные происходят от беннетитовых. С последними примитивные покрытосеменные сходны наличием общих черт в строении древесины, устьица, пыльцы, энтомофилии и т.п. Прародиной покрытосеменных считают районы с сезонно сухим климатом, где они имели наибольшие шансы обживать нарушенные экосистемы благодаря высоким темпам генеративного развития и формирования зародыша. Аналогичными признаками как раз обладали раннемеловые беннетитовые и цикадовые. Есть предположение о появлении признаков, характеризующих цветковые (сосуды в древесине, сетчатое жилкование, завязь, рыльце, двойное оплодотворение), параллельно и порознь у разных групп. Цветковые возникают, когда все эти признаки концентрируются в одной группе. Филогенетические взаимоотношения основных групп растений представлены на рис. 5.
Цветковые растения, постепенно распространяясь, завоевывают обширные пространства. В процессе эволюции покрытосеменных цветок (основной отличающий их орган) претерпевает значительные изменения. Ось цветка — цветоложе — постепенно укорачивается, междоузлия сближаются, спиральное расположение частей цветка переходит в циклическое, происходит процесс уменьшения числа гомологичных частей (олигомеризация).

Рисунок 5- Филогенез основных групп растений (С.В.Мейен)
Первые примитивные энтомофильные цветки привлекали насекомых обилием пыльцы, что одновременно способствовало перекрестному опылению. Преимущество получили те растения, у которых высокая наследственная пластичность потомства, большая вероятность опыления и завязываемости семян. В дальнейшем отбор растений пошел по пути привлечения опылителей с помощью нектара, аромата, окраски и специализации цветков на опыление определенными видами насекомых. Таким путем происходило взаимоприспособление растений и животных по соответствующим признакам. При опылении насекомыми повышается возможность свободного скрещивания растений одного вида, что и служит одной из причин высокой эволюционной пластичности цветковых растений. У цветковых (в отличие от голосеменных) даже деревья представлены множеством разнообразных форм. Цветковые также были приспособлены использовать среду путем быстрого развития и накопления органического вещества.
В кайнозое (начало — 66 млн лет назад) вся Европа была покрыта пышными лесами теплого и умеренного климатов, включающими дуб, березу, сосну, каштан, бук, виноград, орех и др. В это время леса достигали наибольшего распространения на Земле. В тропической флоре этого периода были представлены фикусы, лавровые, гвоздичные, эвкалипты, виноград и др.
В четвертичном периоде кайнозойской эры (2 млн лет назад) увеличилось количество осадков и наступило оледенение значительной части Земли, вызвавшее отступление теплолюбивой третичной растительности на юг (а местами полное ее вымирание), возникновение холодоустойчивых травянистых и кустарниковых растений. На огромных территориях завершается начатая в миоцене смена лесов степью, формируется ксерофитная и эфемерная растительность с выраженной сезонностью в цикле развития, складываются современные фитоценозы.
Таким образом, растительность нашей планеты постоянно менялась, приобретая все более современные черты.
Основные черты эволюции царства растений следующие:
1. Переход от гаплоидности к диплоидности. С диплоидизацией организма снижался эффект проявления неблагоприятных мутаций. У многих водорослей все клетки (кроме зиготы) гаплоидны. У более высокоорганизованных водорослей (бурые и др.) наряду с гаплоидными существуют и диплоидные особи. У мхов преобладает гаплоидное поколение при сравнительно слабом развитии диплоидного. У папоротников преобладает диплоидное поколение, однако и у них гаплоидное поколение (гаметофит) еще представлено самостоятельным образованием, у голосеменных и покрытосеменных наблюдается почти полная редукция гаметофита и переход к диплоидной фазе.
2. Утрата связи процесса полового размножения с наличием капельно-жидкой воды, потеря подвижности мужских гамет, заметная редукция гаметофита и сильное развитие спорофита, переход от наружного оплодотворения к внутреннему, возникновение цветка и двойного оплодотворения.
3. Дифференциациятела с переходом к наземным условиям: деление на корень, стебель и лист, развитие сети проводящей системы, совершенствование покровных, механических и других тканей.
4. Специализация опыления (с помощью насекомых) и распространение семян и плодов животными. Усиление защиты зародыша от неблагоприятных условий: обеспечение пищей, образование покровов и др.
6. Основные пути эволюции животных
Царство животных не менее разнообразно, чем царство растений, а по числу видов животные превосходят растения. Описано около 1 млн 200 тыс. видов животных (из них около 900 тыс. видов — членистоногих, 110 тыс. — моллюсков, 42 тыс. — хордовых животных) и считается, что это может быть лишь половина существующих видов.
Возникновение животных в ископаемых остатках не прослеживается. Первые остатки животных находят в морских отложениях протерозоя, возраст которых превышает 1 млрд лет. Первые многоклеточные животные представлены сразу несколькими типами: губки, кишечнополостные, плеченогие, членистоногие.
В морях кембрийского периода уже существовали все основные типы животных. Облик фауны определяли многочисленные хелицеровые (похожие на современных мечехвостов), губки, кораллы, иглокожие, разнообразные моллюски, плеченогие, трилобиты (рис.6).
После кембрия эволюция животных характеризовалась лишь специализацией и совершенствованием основных типов. Исключение составляют позвоночные, остатки которых обнаружены в ордовике. Это были так называемые щитковые — существа, отдаленно сходные с современными круглоротыми (миноги, миксины), но покрытые со спинной стороны мощно развитыми костными пластинами. Предполагают, что они защищали первых мелких (около 10 см длиной) позвоночных от огромных хищных ракообразных. В теплых и мелководных морях ордовика обитали многочисленные кораллы, значительного развития достигали головоногие моллюски — существа, похожие на современных кальмаров, длиной в несколько метров.
Силурийский период ознаменовался важными событиями не только для растений, но и для животных. Появились животные, дышащие воздухом. Первыми обитателями суши были паукообразные, напоминавшие по строению современных скорпионов. Тем временем в водоемах происходило бурное развитие разнообразных низших позвоночных, прежде всего панцирных рыб.
Предполагается, что первые позвоночные возникли в мелководных пресных водоемах. Постепенно, в течение девона, эти пресноводные формы

Рисунок 6 – Фауна кембрия, ордовика и силура: А-скелет археоциат, Б-древнейшее членистоногое – трилобит, В-скелет коралла, Г-раковина головоногого моллюска, Д-ракоскорпион, Е.З-древнейшие позвоночные – бесчелюстные (Д.К.Беляев и др.,1967)
завоевывают моря и океаны. В девоне же возникают двоякодышащие, кистеперые и лучеперые рыбы. Все они были приспособлены к дыханию в воде. До наших дней дожили некоторые виды двоякодышащих (рис. 7), лучеперые дали начало современным костистым рыбам, а кистеперые — первичным земноводным (стегоцефалам). Стегоцефалы появились в верхнем девоне; примерно в это же время возникает другая чрезвычайно прогрессивная группа животных — насекомые. В развитии линий позвоночных и беспозвоночных проявились две разные тенденции в решении одних и тех же задач. Переход в воздушную среду из водной потребовал укрепления основных несущих органов и всего тела в целом. У позвоночных роль каркаса играет внутренний скелет, у высших форм беспозвоночных — членистоногих — наружный скелет.
Развитие в среде, требовавшей все более сложных поведенческих реакций, решалось в этих двух ветвях древа жизни двумя принципиально разными способами. У насекомых чрезвычайно сложная нервная система, с разбросанными по всему телу огромными и относительно самостоятельными нервными центрами, преобладание врожденных реакций над приобретенными. У позвоночных — развитие огромного головного мозга и преобладание условных рефлексов над безусловными.
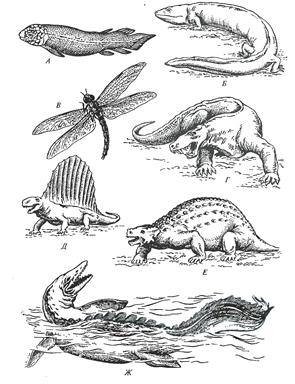
Рисунок 7 – Фауна девона, кембрия и перми: А-двоякодышашая рыба, Б-стегоцефал,
В-стрекозоподобное насекомое, Г-Ж-рептилии (Д.К.Беляев и др.,1967)
В каменноугольном периоде появляются первые пресмыкающиеся, что определило начало активного завоевания суши позвоночными. Рептилии благодаря сухим прочным покровам, яйцам, покрытым твердой скорлупой и не боящимся высыхания, были мало связаны с водоемами. В этом периоде возникают и достигают значительного развития такие древнейшие группы насекомых, как стрекозы и тараканы.
В пермском периоде начинают исчезать стегоцефалы и широко распространяются различные рептилии. От примитивных рептилий из группы цельночерепных в это время развивается ветвь пеликозавров, приведшая несколько позже — через терапсид — к возникновению млекопитающих.
В конце палеозоя происходит значительное иссушение климата. Поэтому бурное развитие претерпевают разнообразные рептилии; до наших дней из триасовых рептилий дожили гаттерия и черепахи. Некоторые рептилии становятся хищниками, другие — растительноядными, третьи — вторично возвращаются в водную среду (рис. 8), обеспечивающую им пищу в виде многочисленных форм костистых рыб и головоногих моллюсков. Однако особенно сильного развития достигают морские рептилии в юре (ихтиозавры, плезиозавры). Тогда же пресмыкающиеся осваивают и воздушную среду — возникают птерозавры, видимо, охотившиеся на многочисленных и крупных насекомых. В триасе от одной из ветвей рептилий возникают птицы; первые птицы причудливо сочетали признаки рептилий и птиц.
В меловом периоде продолжается специализация рептилий: возникают гигантские растительноядные динозавры, встречаются летающие ящеры с размахом крыльев до 20 м. Знаменательные события происходят и в мире насекомых — начинается активная сопряженная эволюция энтомофильных растений и насекомых-опылителей. Происходит процесс вымирания аммонитов, белемнитов, морских ящеров. В связи с похолоданием, вызванном падением астероида, вымирают растительноядные динозавры, а следом — и охотившиеся на них хищные динозавры. Лишь в тропическом поясе сохраняются крупные рептилии (крокодилы). В условиях похолодания исключительные преимущества получают теплокровные животные — птицы и млекопитающие, которые пышно расцветают лишь в следующем периоде — кайнозое.

Рисунок 8 – Пресмыкающиеся мезозоя: А-рогатый динозавр, Б-ихтиозавр, В-летающий хвостатый ящер, Г-бронтозавр, Д, Ж-летающие бесхвостые ящеры, Е-стегозавр,
З- плезиозавр (Д.К.Беляев и др., 1967)
Кайнозой — время расцвета насекомых, птиц и млекопитающих. В конце мезозоя возникают плацентарные млекопитающие. В палеоцене и эоцене от насекомоядных происходят первые хищники. В это же время или несколько позже первые млекопитающие начинают завоевывать море (китообразные, ластоногие, сиреновые). От древних хищных происходят копытные, от насекомоядных обособляется отряд приматов. К концу неогена встречаются уже все современные семейства млекопитающих, на обширных открытых пространствах саванн Африки появляются многочисленные формы обезьян, многие из которых переходят к прямохождению. Одна из групп таких обезьян — австралопитеки — дала ветви, ведущие к роду Homo.
В кайнозое особенно четко проявляются тенденции в развитии самых прогрессивных ветвей древа жизни животных, ведущих к возникновению стайного, стадного образа жизни (что стало ступенькой к последующей социальной эволюции человека).
В четвертичном, или антропогеновом периоде кайнозоя наблюдались резкие изменения климата нашей планеты, связанные с постепенным похолоданием. На этом фоне в средних широтах Северного полушария возникали значительные оледенения суши.
Максимального распространения материковые оледенения достигали во время среднего плейстоцена — около 250 тыс. лет назад. На территории Европы в плейстоцене насчитывается по крайней мере пять таких ледниковых периодов.
Огромное значение для эволюции современной фауны имело то обстоятельство, что одновременно с наступлением ледниковых периодов происходили значительные колебания уровня Мирового океана: в разные периоды этот уровень понижался или повышался на сотни метров сравнительно с современным. При таких колебаниях уровня океана могла обнажаться большая часть материковой отмели Северной Америки и Северной Евразии. Это, в свою очередь, вело к появлению сухопутных «мостов» типа Берингова моста, соединявшего Северную Америку и Северную Евразию, соединению Британских островов с европейским материком и т.п. В Европе 5—6 тыс. лет назад климат был заметно теплее современного. Однако эти изменения климата уже не играли столь значительной роли в изменении видового состава животного мира, какую стал играть Человек, не только уничтоживший многие виды животных и растений (по некоторым подсчетам, человек к середине XX в. уничтожил более 200 видов животных), но и создавший новых домашних животных и ставящий сейчас грандиозную задачу управления эволюционным процессом.
Таким образом, в эволюции животных можно выделить следующие магистральные направления развития адаптаций:
1. Возникновение многоклеточности и все большее дифференцирование всех систем органов.
2. Возникновение твердого скелета (наружного — у членистоногих, внутреннего — у позвоночных).
3. Развитие центральной нервной системы. Два разных и чрезвычайно эффективных эволюционных «решения»: у позвоночных развитие головного мозга, основанного на обучении и условных рефлексах, и возрастание ценности отдельных особей; у насекомых — развитие нервной системы, связанной с наследственным закреплением любого типа реакций по типу инстинктов.
4. Развитие социальности у ряда животных, с разных сторон подходящих к рубежу, отделяющему биологическую эволюцию от социальной. Перешагнуть этот рубеж смогла лишь одна ветвь приматов —
представители рода Homo.
До сих пор остаются гипотетическими конкретные пути возникновения жизни на Земле. Однако возможность возникновения живого на основе процессов химической эволюции через предбиологическую стадию научно
доказана.
Все царства живой природы имеют общее происхождение, связанное с жизнью в первичном океане, но дальнейшие пути развития этих основных стволов древа жизни совершенно различны и по направлениям, и по результатам.
По мере развития новых групп организмов происходит многократное усложнение — биосферы — и эволюция одних групп оказывается тесно связанной с эволюцией других.
Лекция № 15
Тема лекции: Антропогенез
План лекции:
1. Место человека в системе животного мира
2. Непосредственные предшественники человека
3. Основные этапы эволюции рода Homo
4. Пути эволюции Человека разумного
4. 1. Гипотеза широкого моноцентризма
4.2. Особенности ранней стадии эволюции Человека разумного
4.3. Основные достижения в развитии Человека разумного
4.4. Роль труда в процессе возникновения Человека разумного
4.5. Социальная эволюция
Дата добавления: 2017-09-19; просмотров: 469;