Пути эволюции Человека разумного
Существуют две основные точки зрения, касающиеся происхождения человека современного типа. Согласно одной, Н.sapiens возник в нескольких местах планеты из разных предковых форм, принадлежащих к палеоантропам (или даже архантропам). Согласно другой, существовало единое место возникновения человечества из какого-то одного общего предкового ствола. Первая точка зрения — гипотеза полицентризма, вторая — гипотеза моноцентризма. Однако, все четче вырисовывается комплексная, объединяющая все главнейшие аргументы той и другой концепции гипотеза, получившая название гипотезы широкого моноцентризма (Я.Я. Рогинский).
4. 1. Гипотеза широкого моноцентризма
Человек современного типа сложился в окончательном виде где-то в Восточном Средиземноморье и в Передней Азии. Именно здесь находят наиболее выраженные промежуточные между неандертальцами и кроманьонцами костные остатки. Многочисленные промежуточные формы находят также на юго-востоке Европы. В те времена все эти территории были покрыты густыми лесами, населенными разнообразными животными. Здесь, по-видимому, и был совершен последний шаг на пути к Человеку разумному.
На этой стадии был окончательно сформирован облик современного человека. Фигура человека стала более стройной, несколько увеличился рост, лицевая часть черепа стала занимать меньшее место на голове. Емкость черепа, а соответственно и величина головного мозга в целом не изменились, однако в строении мозга произошли значительные перемены: большое развитие получили лобные доли и зоны, связанные с развитием речи и сложной, конструктивной деятельностью.
После этого началось широкое и активное расселение возникших неоантропов по планете. Люди современного типа широко смешивались с жившими сравнительно оседло неандертальцами, что приводило к резкой смене примитивной, неандертальского типа культуры значительно более развитой культурой неоантропов (кроманьонцев).
Вероятно, это было не простое вытеснение (или тем более уничтожение), а различные формы слияния, при которых с генетической точки зрения происходило скрещивание пришлых и местных людей, а с точки зрения развития культуры — начинала безраздельно преобладать новая, прогрессивная культура кроманьонцев.
По данным молекулярной биологии и археологии, современные люди попадали (исходно — из африканского центра происхождения):
- в Переднюю Азию — 73 - 56 тыс. лет назад;
- в Восточную и Центральную Европу — 51 - 39 тыс. лет назад;
- в Северную Америку (по Берингову мосту) — 15-7 тыс. лет назад.
Картина заселения современной мира оказывается очень сложной — периодически наступавший ледник резко сокращал обитаемую территорию в Евразии и Северной Америке. По результатам анализа митохондриальной ДНК прослеживаются, например, два потока заселения Европы перед последним наступлением ледника: 30—35 тыс. лет назад из Северной Азии и 25 тыс. лет назад из Передней Азии. Во время последнего ледникового периода 24—16 тыс. лет назад люди в Европе выжили, по-видимому, только в трех зонах: на Пиренейском полуострове, Балканах и на территории современной Украины. По мтДНК получается, что около 39 тыс. лет назад протоевразийцы попали из Индии - Индокитая в Северную Евразию (предки манси), а через 13 тыс. лет после этого следующая волна расселения человека оставила свои следы в крови современных кетов, нганасан и алтайцев.
Анализ строения митохондриальной ДНК показал, что примерно 150 тысяч лет назад, в плейстоцене, численность исходной популяции кроманьонцев составляла не более 5000 размножающихся пар. Затем, по-видимому, эта популяция разделилась на несколько и каждая из субпопуляций проходила через «бутылочное горлышко» — периоды исключительно низкой численности, когда число размножающихся пар могло быть всего несколько десятков.
Среди нерешенных вопрос о том, когда в ходе эволюции рода Homo возник типичный для современного человека набор из 46 хромосом (у всех человекообразных обезьян — 48 хромосом). Механизм перехода от кариотипа, включающего 2п = 48 хромосом, к кариотипу в 2 п = 46 хромосом понятен. В результате слияния могла возникнуть особь с 47 хромосомами. При скрещивании с 48-хромосомной особью в потомстве появится 50% с 2п = 48 и 50% с 2п = 47. В потомстве особей с 2n = 47 появятся 35% детей с 2п = 46, 25% — 2п = 48 и 50% — 2п = 47. Дальнейшие исследования в помогут определить то место в пространстве и времени, когда возник либо весь род Homo, либо только подвид Н. sapiens sapiens.
4.2. Особенности ранней стадии эволюции Человека разумного
К моменту возникновения кроманьонца род Homo обладал почти всеми признаками, характерными для Человека разумного: прямохождение возникло миллионы лет назад на стадии рамапитеков или даже несколько раньше; начало трудовой деятельности, связанное с использованием передних конечностей, относится к периоду 5 млн лет назад. Примерно тогда же началось использование огня. Непрерывно за последние несколько миллионов лет (и особенно активно за последние несколько сотен тысяч лет) идет наращивание массы головного мозга. Трудовая деятельность вместе со стадным образом жизни приводит к возникновению речи уже у архантропов.
Именно на стадии архантропа возникает членораздельная речь, язык. Об этом свидетельствует появление в лобных долях головного мозга центра Брока (французский анатом), а в височных – центра Вернике (немецкий врач). Эти центры отвечают за способность человека понимать речь. При их выключении у современного человека возникает афазия (человек слышит речь и произносит слова, но не понимает их смысла). У обезьян в этих участках коры расположены центры мимики и жестикуляции, а звуковые сигналы у обезьян принимаются и расшифровываются в подкорке, где анализируются эмоции.
По-видимому, у архантропов произошло «переключение» системы анализа звуковых сигналов с древней подкорки («компьютер 1-ого поколения») на кору (более мощный «компьютер 2-ого поколения»). Это, в свою очередь, стимулировало развитие языка, обеспечило возможность передачи от поколения к поколению сложных трудовых навыков и знаний. Так возникла «вторая сигнальная система» по И.П.Павлову, т.е. способность производить, воспринимать и анализировать отвлечённую вербальную информацию. Наличие этой системы отличает человека от животных, которые всю информацию получают только в виде чувственных образов.
Отдельные группы палеоантропов, сильные своей хитростью (развитый мозг), способностью противостоять самым крупным хищникам (орудия), неблагоприятным природным условиям (жилища, огонь), стали занимать высшие звенья пищевых цепей во всех районах своего обитания. Но пока это все еще отдельные стада, племена, относящиеся друг к другу всегда враждебно (вплоть до каннибализма). Этого требовал естественный отбор — победа любой ценой, в том числе и ценой жизни своих соплеменников или тем более членов соседних племён.
Несомненно, на ранних этапах развития общества существовал
групповой отбор, направленный на возникновение способности ставить превыше всего интересы племени, жертвовать собственной жизнью ради этих интересов. Это и было предпосылкой возникновения социальности, о чем говорил еще Ч. Дарвин: «...те сообщества, которые имели наибольшее число сочувствующих друг другу членов, должны были процветать больше и оставить после себя более многочисленное потомство». Именно отбор по «генам альтруизма» (Джон Холдейн) вывел «человека в люди»! В основе возникновения Человека разумного как вида лежат альтруистические наклонности, определявшие преимущество их обладателей в условиях коллективной жизни.
Необходимо отметить, что со временем простой групповой отбор, как в животном мире, сменился принципиально новым, уже социально мотивированным отбором, связанным с необходимостью передачи накопленного опыта, традиций, объединения усилий отдельных индивидов в процессе труда, охоты и защиты. Произошла грандиозная переоценка ценностей. Раньше неандерталец, видимо, мог спокойно съесть больного, старого или раненого члена своей же семьи. Затем преимущества перед другими получили те племена, которые заботились о стариках и поддерживали физически не таких сильных, но полезных своими умственными способностями особей (мастеров по выделке орудий, хороших следопытов и наблюдателей и т. п.).
С трудом просуществовав около 100 тыс. лет «в тени» биологически прогрессивных неандертальцев, кроманьонцы приобрели какие-то особенности, позволившие им взять верх над своими могучими собратьями — неандертальцами. Примерно 75 тыс. лет назад численность кроманьонцев возросла в десятки раз (достигнув нескольких миллионов), и они на протяжении следующих 15—20 тыс. лет вытеснили неандертальцев и захватили всю планету. Не исключено, что различия касались каких-то особенностей головного мозга. Сравнение 18 тыс. генов человека, шимпанзе и макаки (С. Паабо, 2002) не обнаружило никаких различий в строении ДНК из клеток печени и крови, но очень значительные — в ДНК из клеток головного мозга.
4.3. Основные достижения в развитии Человека разумного
Не рассматривая подробно многих замечательных технических и культурных достижений, характерных для наших предков на первых этапах существования вида Н. sapiens, остановимся на трех моментах.
Первый — небывалое духовное, психическое развитие Человека разумного. Только Человек разумный достиг такого понимания природы, такого уровня самопознания (человек, по философскому определению,— это «материя, познающая самое себя»), который сделал возможным создание искусства (возраст первых наскальных изображений 40—50 тыс. лет).
Не исключено, что это духовное развитие связано с невероятным обогащением половой жизни человека (развитием повышенной сексуальности) по сравнению с тем, что известно для всех других млекопитающих (З.Фрейд, 1929; Д.Моррис, 1969). Об этом говорит и не встречающееся у других видов значительное развитие эрогенных зон у человека, и возникновение женского оргазма — явления неизвестного у обезьян, и полиэстричность человека, и мощное развитие вторичных половых признаков. Не исключено, что уже со стадии архантропов, и, несомненно,— со времени кроманьонцев, половое влечение, любовь в ее физическом и психологическом смысле во многом определяет высокий уровень социальности и развитие человеческого общества.
Вторым величайшим достижением эволюции Человека разумного были открытия, приведшие к неолитической революции — приручению животных и окультуриванию растений (около 11 тыс. лет назад). Эти события были, пожалуй, наиболее крупными на пути овладения Н. sapiens окружающей средой. До этого человек был полностью зависим от среды при добыче пищи и одежды. Теперь зависимость стала значительно меньшей.
Масштабы неолитической революции ныне часто недооцениваются. Можно напомнить, что практически все домашние животные и культурные растения созданы нашими далекими предками много тысяч лет назад. В культуру были введены и одомашнены множество видов, потом «утерянных» в этом качестве. Среди домашних животных в Европе уже в историческое время были хорьки и ласки, а в Средней Азии — гепарды, в Китае — бакланы и т.д.
Важным следствием неолитической революции стало появление у человека времени, свободного от добывания пищи. Появление такого свободного времени должно было быть мощным стимулом для развития культуры, искусства, ремесел, религии.
Третьим крупнейшим этапом в истории современного Человека была научно-техническая революция, в результате которой человек приобрел власть над природой (в последние 2 тыс. лет и особенно в последние 3—4 столетия). Техническая мощь человечества ныне такова, что оно может по своему желанию менять условия существования в крупных районах нашей планеты, обеспечивать условия для жизни в районах, прежде совершенно недоступных (Арктика и Антарктика), в воздухе, на воде, наконец, в ближнем Космосе.
Выделенные важнейшие вехи в развитии Homo sapiens не связаны, как показывают антропологические и археологические данные, с изменениями физического облика человека за последние десятки тысяч лет. Различия между первыми кроманьонцами и современными людьми намного меньше, чем различия между отдельными современными этническими группами и расами. Отсутствие уловимых тенденций в изменении строения тела человека за несколько последних десятков тысяч лет косвенно показывает, что эволюция Человека вышла из-под ведущего контроля биологических факторов и направляется действием иных — социальных сил.
4.4. Роль труда в процессе возникновения Человека разумного
Главные особенности человека: развитие центральной нервной системы; руки как органа, способного производить сотни разнообразных движений, недоступных обезьянам; речь как средство общения людей и создание общества как такового — все это результат процесса труда, производства. Напомним, что возникновение рода Человека, первым изученным представителем которого мы считаем Человека умелого (Н. habilis), связано именно с трудовой деятельностью. Не просто использование палки, камня, кости как орудия (такое использование предметов характерно и для современных обезьян и, конечно, было широко распространено среди австралопитеков), но именно производство различных орудий — вот грань, отделяющая человека от человекоподобных предков. Впервые решающую роль труда в происхождении человека отметил Фридрих Энгельс в книге «Роль труда в процессе превращения обезьяны в человека» (1876). Все дальнейшее развитие Человека связано с совершенствованием процесса производства, в конце концов приведшего Человека к власти над природой.
4.5. Социальная (культурная) эволюция
Биологическая эволюция в основном закончилась около 40 тыс. лет назад, когда окончательно сформировался неоантроп. До этого основным механизмом антропогенеза был естественный отбор на лучшую
орудийную деятельность. После этого биологическая эволюция сменилась социальной, а главным механизмом антропогенеза стал естественный отбор на способность к накоплению, передаче и анализу знаний, накопленных предыдущими поколениями.
Тесно связана с развитием этой способности эволюция материальных носителей информации. На ранних этапах развития человека накопленные знания и навыки передавались путём копирования физических действий, затем – вербально. Сами предметы материальной культуры (орудия, украшения, одежда, жилища и т.д.) можно считать первыми примитивными носителями информации. Далее появились рисунки как способ накапливать и сохранять знания и опыт. Затем появляется письменность. Настоящая революция – книгопечатание. И, наконец, -с середины 50-х гг. ХХ в. – новая революция -создание и широкое использование ЭВМ.
Простое перечисление показывает, что с каждым новым этапом
ёмкость, продолжительность хранения и тираж носителей информации увеличивались, открытия отделялись от их создателей, знание становилось отвлеченным иболее доступным. Всё это способствовало ускорению социальной эволюции человека и всё большему его отдалению от мира природы.
По скелетным остаткам можно воссоздать внешний облик и сравнить между собой людей, живших 40—30 тыс. лет назад, 10—5 тыс. лет назад, 500 лет назад, и современных людей. Существенных различий в строении тела между этими группами людей нет. В то же время различия несомненно есть в культуре. Современный человек знает о свойствах окружающего его мира неизмеримо больше, чем его прародители, он ставит и обсуждает проблемы, представление о которых отсутствовало у предыдущих поколений. Все знания передаются следующим поколениям не автоматически, а с помощью обучения. Так, путем вертикальной передачи приобретенных знаний осуществляется социальное развитие, закономерности которого оказываются совершенно иными, нежели закономерности биологической эволюции. Одновременно с совершенствованием способности к передаче утилитарных навыков человек развивал свой культурный мир, создавал не только орудия, но и мифы,религии, наукииискусства.
Некоторые этапы социальной (культурной) эволюции Homo:
• копье — 40 тыс. лет назад;
• украшения — 20 тыс. лет назад;
•скульптурные изображения женщины, наскальные и накостные рисунки мамонтов, львов, туров, носорогов — 35—32 тыс. лет назад;
•музыкальный инструмент (флейта) — 25 тыс. лет назад;
• лук и стрелы — 18 тыс. лет назад;
• расцвет наскальной живописи — 17—15 тыс. лет назад;
• глиняный кирпич — 12 тыс. лет назад;
• глиняный сосуд — 8 тыс. лет назад;
• колесо-диск — 6,5 тыс. лет назад
• гончарный круг — 6 тыс. лет назад;
• пиктография — 5,5 тыс. лет назад;
• папирус — 4,7 тыс. лет назад;
• колесо со спицами — 3,5 тыс. лет назад;
• алфавит с гласными — 3,2 тыс. лет назад.
Несомненно, социальная эволюция возникла на основе биологической. Какое-то значительное время оба типа эволюции сосуществовали, оказывая влияние на все развитие рода Homo. При этом влияние биологической эволюции уменьшалось, а социальной — увеличивалось.
Возникновение трудовой деятельности, производства орудий труда явилось качественным скачком, поворотным моментом от истории биологической (филогенеза) к истории социальной.
Своеобразие эволюции рода Homo состоит в том, что биологические эволюционные факторы постепенно теряют свое ведущее значение, уступая социальным факторам. Возникший в процессе эволюции как часть животного мира Человек разумный в результате общественно-исторического развития настолько выделился из природы, что приобрел власть над ней. Насколько разумно и дальновидно сумеет он использовать эту власть — вопрос будущего.
Лекция № 16
Тема лекции: Расогенез
План лекции:
1. Классификации рас
2. Гипотезы о происхождении рас
3. Этапы расообразования
4. Факторы расогенеза
5. Расизм и его критика
6. Возможные пути эволюции человека в будущем
Классификации рас
Раса — это исторически сложившаяся группировка (группа популяций) людей, характеризующихся морфо-физиологическим сходством и происходящих из определённого географического региона. Развиваясь под влиянием исторических факторов и принадлежа к одному виду (H.sapiens), раса отличается от народа, или этноса, который, обладая определенной территорией расселения, может состоять из представителей нескольких рас, или, наоборот, к одной и той же расе может принадлежать ряд народов и носителей многих языков.
В более широком понимании термин расаиспользуется по отношению к растениям и животным и обозначает наследственную внутривидовую группировку, меньшую по рангу, чем подвид.
О существовании рас люди знали еще до нашей эры. Тогда же были предприняты и первые попытки объяснить их происхождение. Например, в мифах древних греков происхождение людей с черной кожей объяснялось неосторожностью Фаэтона (сына бога Гелиоса), который на солнечной колеснице так приблизился к Земле, что обжег стоявших на ней белых людей. Греческие философы в объяснениях причин возникновения рас большое значение придавали климату. В соответствии с Ветхим Заветом, родоначальниками белой, черной и желтой рас были сыновья Ноя — любимый богом Яфет, Сим и проклятый богом Хам (соответственно).
Стремления систематизировать представления о физических типах народов, населяющих земной шар, датируются XVII в., когда, опираясь на различия людей в строении лица, цвета кожи, волос, глаз, а также особенности языка и культурных традиций, французский врач Ф. Бернье впервые в 1684 г. ввёл понятие «раса» и классифицировал человечество на три расы — европеоидную, негроидную и монголоидную. Сходную классификацию предлагал К. Линней, который, признавая человечество в качестве единого вида, выделял дополнительную (четвертую) расу — лапландскую (население северных районов Швеции и Финляндии).
Француз Ж. Блюменбах в 1775 г. разделил род человеческий на основе цвета кожи на пять рас — кавказскую (белую), монгольскую (желтую), эфиопскую (черную), американскую (красную) и малайскую (коричневую), а У. Бойд в 1953 г. на основе результатов изучения антигенов крови поделил человечество на пять рас (групп):
1. Европейская, включая лопарей, южных европейцев и северных африканцев.
2. Африканская группа.
3. Азиатская группа, включая жителей индийского субконтинента.
4. Американская группа, включая всех аборигенов.
5. Тихоокеанская группа (меланезийцы, полинезийцы, австралоиды).
В практическом плане очень популярна классификация рода человеческого, делящая его на три большие расы: европеоидов, негроидов и монголоидов.
В рамках каждой большой расы выделяют отдельные антропологические типы с устойчивыми комплексами признаков, называющиеся малыми (локальными) расами.
Существует три основных подхода к классификации рас: без учета их происхождения, с учетом происхождения и родства и на основе популяционной концепции. В соответствии с первым подходом, три большие расы включают в себя 22 малые, причем между большими расами располагаются по две переходные малые. Схема расовой классификации изображается при этом в виде круга (рис.1).
Несмотря на то, что при такой классификации не учитывается происхождение рас, само существование малых переходных рас, сочетающих в себе одновременно признаки двух больших рас (эфиопская, южносибирская, уральская и т.д.), свидетельствует, с одной стороны, о динамизме расовых комплексов признаков, а с другой — об условности членения человечества даже на большие расы.
Переходные малые расы совмещают в себе не только морфологические признаки в соответствии с их промежуточным положением, но являются промежуточными и в генетическом отношении. Из этого следует, что они либо гибридогенны, либо сохранили в своей организации более
древние черты, характерные для этапа существования человечества, предшествующего формированию больших рас.
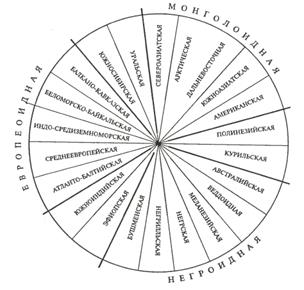
Рисунок 1- Классификация малых рас человека без учёта их происхождения (Я.Я.Рогинский, М.Г.Левин, 1978)
Классификация с учетом происхождения рас изображается в виде эволюционного древа с коротким общим стволом и расходящимися от него ветвями (рис.2).

Рисунок 2 – Классификация рас с учётом их происхождения (В.Н.Ярыгин и др., 2004)
В основе таких классификаций лежит обнаружение черт архаизма и эволюционной продвинутости отдельных рас, в соответствии с чем разные большие и малые расы занимают разное положение на ветвях такого древа. Выявление архаичных (сходство с обезьянами) и прогрессивных черт среди морфологических признаков носит субъективный характер, благодаря чему схемы расовых классификаций такого рода очень многообразны. Но самым большим недостатком подходак классификации рас исходя из их происхождения является попытка расположить расы на разных уровнях эволюционного древа, т.е. признание их биологической неравноценности.
Кроме того, данные палеоантропологических исследований показывают, что вплоть до верхнего палеолита на территориях, заселённых людьми, практически нигде не сформировались расовые типы человека, с которыми были бы генетически связаны современные большие расы. Это подтверждает анализ верхнепалеолитических находок скелетов людей современного физического типа из сунгирских погребений (Россия, под Воронежем), живших приблизительно 26 тыс. лет назад. Все черепа, принадлежащие им, характеризуются мозаичным сочетанием расовых признаков и не могут быть отнесены ни к одной из современных рас.
Только более поздние мезолитические находки свидетельствуют об окончательном формировании у человека расовых признаков. Так, известны мезолитические черепа с территории Северной Африки возрастом 10—8 тыс. лет с явными признаками не просто негроидной, а сразу малой эфиопской расы. Сходные данные получены на территории Европы и в других регионах. Все это указывает на то, что процесс формирования расовых признаков шёл параллельно в разных регионах и завершился 10—8 тыс. лет назад формированием малыхрас на основе племён, обитающих на одной территории и близких по культуре. Большие же расы, вероятно, являются условной, сборной группой.
Это заключение хорошо согласуется с популяционнойконцепциейрас.Суть ее заключается в следующем. Если принять, что большие расы человека представляют собой огромные популяции, то малые расы —
субпопуляции больших, внутри которых имеются ещё более мелкие естественные общности людей – конкретные этносы (нации, народности). Если предположить при этом, намеренно упрощая ситуацию, что этносы не разделяются на элементарные популяции (демы, изоляты), и считать их просто состоящими из конкретных индивидов, то получится сложная структура, включающая в себя четыре уровня иерархии (рис. 3).
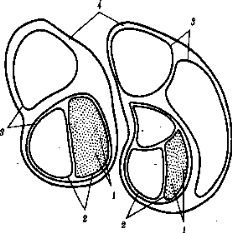
Рисунок 3- Популяционная концепция рас (схема): 4 – большие расы, 3 – малые расы,
2 – этносы, 1 – индивидуумы (В.Н.Ярыгин и др., 2004)
Рассматривая расогенез в качестве процесса, продолжающегося и в настоящее время, Ф.Г. Добржанский (1962) классифицировал человечество на 34 расы, а именно:
1. Северо-западная европейская — жители Скандинавии, северной части ФРГ, Северной Франции, Великобритании и Ирландии.
2. Северо-восточная европейская — жители Польши, Европейской части России, Украины, Белоруссии, большинство существующих популяций в Сибири.
3. Альпийская — жители территорий, простирающихся от Центральной Франции, южной части ФРГ, Швейцарии, Северной Италии до побережья Черного моря.
4. Средиземноморская — популяции по обе стороны Средиземноморья, от Танжера до Дарданелл, Аравии, Турции, Ирака.
5. Хинду — жители Индии, Пакистана.
6. Тюркская — жители Туркестана, Западного Китая.
7. Тибетская — жители Тибета.
8. Северо-китайская — жители Северного и Центрального Китая, Маньчжурии.
9. Классическая монголоидная — жители Сибири, Монголии, Кореи, Японии.
10. Эскимосская — жители арктической Азии и Америки.
11. Юго-восточная азиатская — жители Южного Китая до Таиланда, Бирмы, Малайзии и Индонезии.
12. Айну — популяция аборигенов в Северной Японии.
13. Лопари (саами) — аборигены арктической Скандинавии и Финляндии.
14. Североамериканская индейская — туземные популяции Канады и США.
15. Центрально-американская индейская — обитатели территорий, простирающихся от юго-западных районов Северной Америки через Центральную Америку до Боливии.
16. Южноамериканская индейская — популяции Перу, Боливии и Чили, занимающиеся главным образом сельским хозяйством.
17. Фужиан - аборигены юга Южной Америки, не занимающиеся сельским хозяйством.
18.Восточно-африканская - популяции Восточной Африки, Эфиопии, части Судана.
19. Суданская — популяции большей части Судана.
20. Лесная негроидная — популяции, обитающие в лесах Западной Африки и вдоль большей части реки Конго.
21. Банту — аборигены Южной Африки и части Восточной Африки.
22. Бушмены и готтентоты — аборигены, населяющие Южную Африку.
23. Африканские пигмеи - популяции малорослых людей, обитающих в лесах Экваториальной Африки.
24. Дравидианы — аборигены Южной Индии и Цейлона.
25. Негрито — популяция малорослых людей с вьющимися волосами на территории от Филиппин до Андаманса, Малайи и Новой Гвинеи.
26. Меланезийская папуасская — аборигены Новой Гвинеи до Фиджи.
27. Мураджиан — популяция аборигенов Юго-Восточной Австралии.
28. Карпентариан — популяция аборигенов Северной и Центральной Австралии.
29. Микронезийская — популяции островов западной части Тихого океана.
30. Полинезийская - популяции островов центральной и восточной части Тихого океана.
31.Неогавайская — популяция, недавно возникшая на Гавайских островах.
32. Ладино — популяция, недавно возникшая в Центральной и Южной Америке.
33. Североамериканская цветная — негритянская популяция в США.
34. Южноафриканская цветная — негритянская популяция Южной Африки.
Рассмотрим основные отличительные особенности рас.
Европеоиды светлокожи, для них характерны прямые или волнистые светло-русые или темно-русые мягкие или средней жесткости волосы, серые, серо-зеленые, голубые и каре-зеленые широко открытые глаза, умеренно развитый подбородок, широкий таз, узкий и сильно выступающий нос, нетолстые губы и довольно обильный волосяной покров тела и лица. Для женщин этой расы характерны полусферические груди и выпуклые ягодицы. Люди, принадлежащие к этой расе, занимают большую часть Европы, а также прилегающие к ней области.
Негроиды темнокожи, для них характерны курчавые или шерстистые темные волосы, толстые губы, очень широкий и плоский нос с продольными ноздрями, очень крупные зубы, карие глаза, длинная голова, редкая растительность на лице и теле, узкий таз, большие ступни. Для женщин характерны груди конической формы и мало выпуклые ягодицы. Люди, принадлежащие к этой расе, занимают практически весь экваториальный пояс от Африки до островов Тихого океана. К этой расе принадлежит население Африки, а также негрито (пигмеи), океанические негроиды (меланезийцы), южно-африканские бушмены и готтентоты.
Монголоиды смуглокожи, обладают желтой или желто-коричневой кожей. Для них характерны прямые жесткие иссиня-черные волосы, плоское скуластое лицо, узкие и слегка раскосые карие глаза со складкой верхнего века (третье веко, или эпикантус) во внутреннем углу глаза, плоский и довольно широкий нос, редкая растительность на лице и теле. Люди, принадлежащие к этой расе, занимают Восточную Сибирь и Монголию, Дальний Восток, Центральную и Юго-Восточную Азию. Видоизменённую монголоидную расу представляют индонезийцы и американские индейцы.
Часто в виде отдельной расы выделяют австралоидов, которые почти так же темнокожи (их кожа имеет шоколадный цвет), как и негроиды, но для них характерны темные волнистые волосы, крупная голова и массивное лицо с очень широким и плоским носом, выступающим подбородком, значительный рост волос на лице и теле. Австралоиды являются аборигенами Австралии. Однако австралоидов часто считают негроидами.
Иногда как отдельную расу выделяют америндов (американских индейцев), для которых характерны смуглая кожа, скуластое лицо, достаточно выступающий нос и эпикантус, иссиня-черные волосы. Однако чаще америндов относят к монголоидам.
Данные о распределении групп крови и типов пальцевых узоров в популяциях, принадлежащих к разным расам, представлены в табл.1 и 2.
Таблица 1 - Распределение групп крови в разных популяциях, %
| Популяция | А | АВ | В | Rh | |
| Европеоиды | 5-40 | 1-37 | 4-18 | 45-75 | 25-46 |
| Негроиды | 8-30 | 1-8 | 10-20 | 52-70 | 4-29 |
| Монголоиды | 0-45 | 0-5 | 16-25 | 39-68 | 0-5 |
| Американские индейцы | 0-20 | около 0 | 0-4 | 68-100 | около 0 |
Правильное понимание процессов расогенеза зависит от подходов к определению сущности и классификации рас. Длительное время господствовал так называемый типологический подход, в соответствии с которым определение расы проводили на основе стереотипов, якобы отражающих все признаки расы. Поэтому, руководствуясь характеристиками отдельных индивидов, считали, что между расами существуют абсолютные различия, что не соответствует действительности.
Таблица 2 - Распределение типов пальцевых узоров в разных популяциях
| Популяция | Встречаемость пальцевых узоров, % | ||
| дуга | петля | завиток | |
| Европеоиды | 0-9 | 63-76 | 20-42 |
| Негроиды | 3-12 | 53-73 | 20-40 |
| Бушмены | 13-16 | 66-68 | 15-21 |
| Китайцы, японцы и родственные популяции | 1-5 | 43-56 | 44-54 |
| Американские индейцы | 46-61 | 35-57 | |
| Австралийские аборигены | 0-1 | 28-46 | 52-73 |
| Микронезийцы | 49-50 |
Научно обоснованным подходом к классификации рас является
популяционный. Эффективность классификаций рас зависит от количества используемых признаков, поскольку чем больше признаков для проведения различий между расами, тем более эффективной оказывается классификация. Поскольку расы являются популяциями, то, изучая какую-либо расу, описывают фактически большую группу людей, но не отдельного индивида. Поскольку в пределах каждой расы имеются громадные вариации наследственных характеристик, то невозможно найти абсолютные критерии, на основе которых можно было бы подтвердить или исключить принадлежность индивидуума к той или иной расе.
Расы должны рассматриваться как группы, но не как индивиды. Например, сравнение черепных костей шведов и русских показало, что между этими национальными группами имеются существенные различия, но они являются средними различиями, а между индивидуумами обеих групп имеется перекрываемость, т.е. зияние (хиатус)отсутствует. Таким образом, определение расовых различий по популярным стереотипам (типологически) является неправильным, поскольку абсолютных различий между расовыми группами не существует.
Дата добавления: 2017-09-19; просмотров: 361;