Изображение А- и В-форм
Двойной спирали ДНК
Третичная структура ДНК. У всех живых организмов двухспиральные молекулы ДНК плотно упакованы с образованием сложных трехмерных структур.
Двухцепочечные ДНК прокариот, имеющие кольцевую ковалентно-замкнутую форму, образуют левые (—) суперспирали. Третичная структура ДНК эукариотических клеток также образуется путем суперспирализации, но не свободной ДНК, а ее комплексов с белками хромосом (белки-гистоны классов Н1, Н2, Н3, Н4 и Н5) (рисунок 2.22).
 Рисунок 2.22 – Схематическое изображение третичной структуры ДНК
Рисунок 2.22 – Схематическое изображение третичной структуры ДНК
|
В пространственной организации хромосом можно выделить несколько уровней. Первый уровень – нуклеосомный. В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм и укорачивается примерно в 7 раз.
Вторым уровнем пространственной организации хромосом является образование из нуклеосомной нити хроматиновой фибриллы диаметром 20— 30 нм (уменьшение линейных размеров ДНК еще в 6—7 раз).
Третичный уровень организации хромосом обусловлен укладкой хроматиновой фибриллы в петли. В образовании петель принимают участие негистоновые белки. Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 пар нуклеотидов. В результате такой упаковки линейные размеры ДНК уменьшаются примерно в 200 раз. Петлеобразная доменная организация ДНК, называемая интерфазной хромонемой, может подвергаться дальнейшей компактизации, степень которой меняется в зависимости от фазы клеточного цикла.
СТРОЕНИЕ РНК
В цитоплазме клеток содержатся три основных функциональных вида РНК: матричные РНК (мРНК), выполняющие функции матриц белкового синтеза; рибосомные РНК (рРНК), выполняющие роль структурных компонентов рибосом; и транспортные РНК (тРНК), участвующие в трансляции (переводе) информации мРНК в последовательность аминокислот молекулы белка.
В ядре клеток обнаруживают ядерную РНК, составляющую от 4 до 10% от суммарной клеточной РНК. Основная масса ядерной РНК представлена высокомолекулярными предшественниками рибосомных и транспортных РНК. Предшественники высокомолекулярных рРНК (28 S, 18 S и 5 S РНК) в основном локализуются в ядрышке.
РНК является основным генетическим материалом у некоторых вирусов животных и растений (геномные РНК). Для большинства РНК вирусов характерна обратная транскрипция их РНК генома, направляемая обратной транскриптазой.
Все рибонуклеиновые кислоты представляют собой полимеры рибонуклеотидов, соединенных, как в молекуле ДНК, 3',5'-фосфорнодиэфирными связями. В отличие от ДНК, имеющей двухцепочечную структуру, РНК представляет собой одноцепочечные линейные полимерные молекулы.
Строение мРНК. мРНК — наиболее гетерогенный в отношении размеров и стабильности класс РНК. Содержание мРНК в клетках составляет 2—6% от общего количества РНК. мРНК состоят из участков — цистронов, определяющих последовательность аминокислот в кодируемых ими белках.
Строение тРНК. Транспортные РНК выполняют функции посредников (адаптеров) в ходе трансляции мРНК. На их долю приходится примерно 15% суммарной клеточной РНК. Каждой из 20 протеиногенных аминокислот соответствует своя тРНК. Для некоторых аминокислот, кодируемых двумя и более кодонами, существуют несколько тРНК.
тРНК представляют собой сравнительно небольшие одноцепочечные молекулы, состоящие из 70—93 нуклеотидов. Их молекулярная масса составляет (2,4—3,1) .104 кДа.
Вторичная структура тРНК формируется за счет образования максимального числа водородных связей между внутримолекулярными комплементарными парами азотистых оснований. В результате образования этих связей полинуклеотидная цепь тРНК закручивается с образованием спирализованных ветвей, заканчивающихся петлями из неспаренных нуклеотидов. Пространственное изображение вторичных структур всех тРНК имеет форму клеверного листа (рисунок 2.23).
В «клеверном листе» различают четыре обязательные ветви, более длинные тРНК, кроме того, содержат короткую пятую (дополнительную) ветвь. Адапторную функцию тРНК обеспечивают акцепторная ветвь, к 3'-концу которой присоединяется эфирной связью аминокислотный остаток, и противостоящая акцепторной ветви антикодоновая ветвь, на вершине которой находится петля, содержащая антикодон. Антикодон представляет собой специфический триплет нуклеотидов, который комплементарен в антипараллельном направлении кодону мРНК, кодирующему соответствующую аминокислоту.
Т-Ветвь, несущая петлю псевдоуридина (ТyС-петлю), обеспечивает взаимодействие тРНК с рибосомами.
Д-ветвь, несущая дегидроуридиновую петлю, обеспечивает взаимодействие тРНК с соответствующей аминоацил-тРНК-синтетазой.
 Рисунок 2.23 - Вторичная
структура тРНК
Рисунок 2.23 - Вторичная
структура тРНК
|
 Функции пятой дополнительной ветви пока мало исследованы, вероятнее всего она уравнивает длину разных молекул тРНК.
Функции пятой дополнительной ветви пока мало исследованы, вероятнее всего она уравнивает длину разных молекул тРНК.
 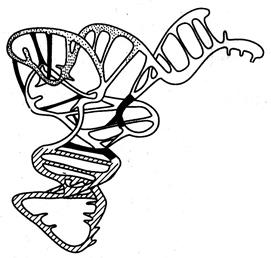 Рисунок 2.24 - Третичная структура тРНК (по А.С. Спирину)
Рисунок 2.24 - Третичная структура тРНК (по А.С. Спирину)
|
Третичная структура тРНК очень компактна и образуется путем сближения отдельных ветвей клеверного листа за счет дополнительных водородных связей с образованием L-образной структуры «локтевого сгиба» (рисунок 2.24). При этом акцепторное плечо, связывающее аминокислоту, оказывается расположенным на одном конце молекулы, а антикодон — на другом.
Строение рРНК и рибосом. Рибосомные РНК формируют основу, с которой связываются специфические белки при образовании рибосом. Рибосомы — это нуклеопротеиновые органеллы, обеспечивающие синтез белка на мРНК. Число рибосом в клетке очень велико: от 104 у прокариот до 106 у эукариот. Локализуются рибосомы главным образом в цитоплазме, у эукариот, кроме того, в ядрышке, в матриксе митохондрий и строме хлоропластов. Рибосомы состоят из двух субчастиц: большой и малой. По размерам и молекулярной массе все изученные рибосомы делят на 3 группы — 70S рибосомы прокариот (S-коэффициент седиментации), состоящие из малой 30S и большой 50S субчастиц; 80S рибосомы эукариот, состоящие из 40S малой и 60S большой субчастиц.
Малая субчастица 80S рибосом образована одной молекулой рРНК (18S) и 33 молекулами различных белков. Большая субчастица образована тремя молекулами рРНК (5S, 5,8S и 28S) и примерно 50 белками.
Вторичная структура рРНК образуется за счет коротких двуспиральных участков молекулы — шпилек (около 2/3 рРНК), 1/3 — представлена однотяжевыми участками, богатыми пуриновыми нуклеотидами.
Дата добавления: 2017-08-01; просмотров: 1009;