Глицерофосфолипиды.
Общим структурным фрагментом всех глицерофосфолипидов является фосфатидная кислота (1,2-диацил,3-фосфоглицерол):
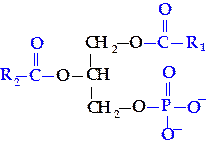
Молекулы фосфолипидов имеют гидрофобную часть, образованную радикалами жирных кислот, и гидрофильную — остатки фосфорной кислоты, аминокислот, аминоспиртов.
Фосфатидилхолин (лецитин). В своем составе содержит аминоспирт холин (гидроксид 3-гидроксиэтилтриметиламмония). В зависимости от того, с каким атомом углерода глицерола связана фосфорная кислота (a— в крайнем положении, b — в срединном), различают два типа фосфатидилхолинов (a и b).
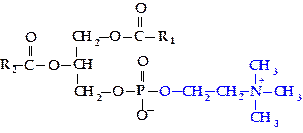
|
Фосфатидилхолины (ФХ) широко распространены в клетках, особенно мозговой ткани человека и животных; в растениях они встречаются в соевых бобах, семенах подсолнечника, зародышах пшеницы.
Фосфатидилэтаноламин (кефалин). В состав фосфатидилэтаноламинов (ФЭА) вместо холина входит азотистое основание этаноламин.
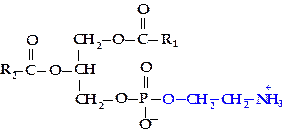
|
ФХ и ФЭА являются главными липидными компонентами мембран клеток.
В результате гидролиза при действии специфичного фермента — фосфолипазы А2 в ФХ или ФЭА возможно отщепление остатка жирной кислоты с образованием лизоформ: лизофосфатидилхолина (ЛФХ) и лизофосфатидилэтаноламина (ЛФЭА):
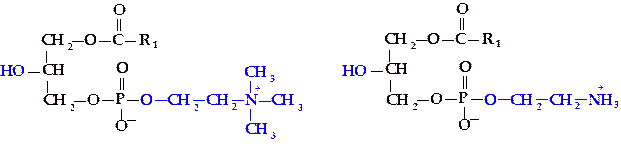
|
Фосфатидилсерин. В молекуле фосфатидилсерина полярной группой является остаток аминокислоты серина.
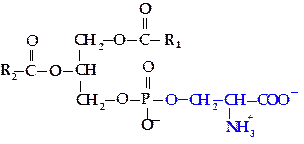
|
Фосфатидилсерины (ФС) распространены менее широко, чем ФХ и ФЭА. Значение ФС определяется тем, что он является предшественником в синтезе двух других групп.
Плазмалогены. Известны также глицерофосфолипиды, которые в отличие от приведенных выше — ФХ, ФЭА, ФС — отличаются тем, что вместо остатка кислоты при атоме углерода С, они содержат a-, b-ненасыщенный спирт, образующий простую эфирную связь с гидроксильной группой глицерола.
При гидролизе этой эфирной связи образуется альдегид соответствующего спирта. Отсюда название группы — фосфатидали: фосфатидальэтаноламин, фосфатидальхолин, фосфатидальсерин.
 фосфатидальэтаноламин (плазмалоген)
фосфатидальэтаноламин (плазмалоген)
|
На долю плазмалогенов приходится до 10% фосфолипидов мозга и мышечной ткани. Они обнаружены также в эритроцитах, тканях некоторых беспозвоночных (до 25%), входят в состав бактериальных мембран, но практически не встречаются в растениях.
Фосфатидилинозитол. В отличие от других групп глицерофосфолипидов в состав фосфатидилинозитола вместо азотсодержащих соединений входит шестиуглеродный циклический спирт инозитол, представленный одним из его стереоизомеров — моноинозитолом.
Фосфатидилинозитол входит в состав клеточных мембран животных, высших растений, различных микроорганизмов, особенно высоко его содержание в миелиновых оболочках нервных волокон.
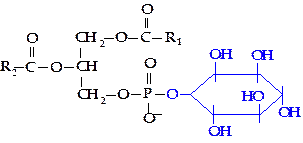
|
Важное значение имеют фосфорилированные производные фосфатидилинозитолов.
Фосфатидилглицерол. Так же как фосфатидилинозитол, фосфотидилглицерол не содержит азотсодержащего соединения. В этих фосфолипидах полярной группой служит еще одна молекула глицерола.
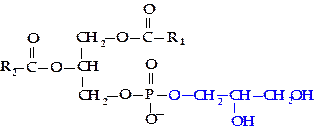
|
Фосфатидилглицеролы и их аминокислотные производные в значительном количестве содержатся в бактериальных мембранах, а также хлоропластах растений.
Кардиолипин. Этот фосфолипид можно рассматривать как производное фосфатидилглицерола, в котором 3-гидроксигруппа второго остатка молекулы глицерола этерефицирована молекулой фосфатидной кислоты.

Кардиолипин локализован почти исключительно в митохондриях и играет важную роль в структурной организации и функционировании дыхательных комплексов. Кардиолипин является также обязательным компонентом бактериальных клеточных мембран и хлоропластов растений.
Сфингофосфолипиды.
В составе соединений этого класса глицерола нет. Они являются производными сложного ненасыщенного дигидроксиаминоспирта — сфингозина или его насыщенного аналога — дигидросфингозина.
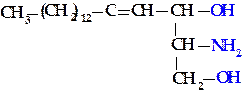
|
При ацилировании Н2N-группы сфингозина жирной кислотой образуется соединение — церамид, фосфохолиновое производное которого называется сфингомиелином. Сфингомиелины являются наиболее распространенными сфинголипидами, находятся в мембранах животных и растительных клеток. Особенно богата ими нервная ткань. Сфингомиелины обнаружены также в ткани почек, печени и других органов, входят в состав липидов крови.
Гликолипиды (гликосфинголипиды). Липиды этого класса, подобно сфингомиелинам, являются производными церамидов, спиртовая группа которых гликозилирована остатками одного или нескольких углеводов. В зависимости от состава углеводного компонента их делят на цереброзиды и ганглиозиды.
Цереброзиды – это церамидомоносахариды, к ним относятся галактозил-церамиды и глюкозилцерамиды. Гликозидная связь с углеводом имеет обычно Р-конфигурацию.
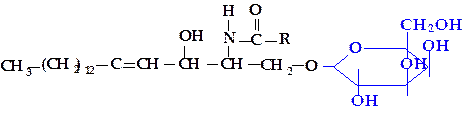
|
Галактозилцерамид является основным гликолипидом мозговой и нервной тканей, содержит различные жирные кислоты, в том числе цереброновую (С24), являющуюся гидроксикислотой. В результате сульфатирования галактозилцерамида он превращается в сульфатид.
Ганглиозиды представляют собой более сложные сфинголипиды, чем цереброзиды. В их состав входят сфингозин, жирная кислота, один или несколько углеводов (D-глюкоза, D-галактоза, N-ацетилглюкозамин, N-ацетилгалактозамин, N-ацетилнейраминовая кислота и др.).
Ганглиозиды в больших количествах находятся в нервной ткани. В сером веществе мозга ганглиозиды составляют около 6% мембранных липидов. Ганглиозиды выполняют рецепторные функции. Они активно участвуют в контроле и регуляции межклеточных контактов, рецепции.
Стероиды являются производными циклопентанпергидрофенантрена, содержащего три нелинейно конденсированных насыщенных циклогексановых и одно циклопентановое кольцо.
К стероидам относится большое количество биологически важных соединений: собственно стеролы (или стерины), витамины группы D, поло-
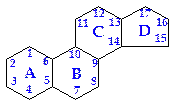 циклопентанпергидрофенантрен
циклопентанпергидрофенантрен
|
вые гормоны, гормоны коры надпочечников, зоо- и фитоэкдистероидные гормоны, сердечные гликозиды, растительные сапонины и алкалоиды, некоторые яды и др.
Классификация стероидов основана на структуре заместителя у С-17 углеродного атома (длина углеродной цепи), числе и положении гидроксигрупп, степени ненасыщенности.
Наиболее известный среди стеролов — холестерол, содержащийся почти во всех тканях организма. Особенно много его в центральной и периферической нервной системе, подкожном жире, почках и др. Холестерол является одним из главных компонентов цитоплазматической мембраны, а также липопротеинов плазмы крови. В липопротеиновых фракциях крови примерно только одна треть его находится в виде спирта, а две трети — в форме эфиров жирных кислот (холестеридов):
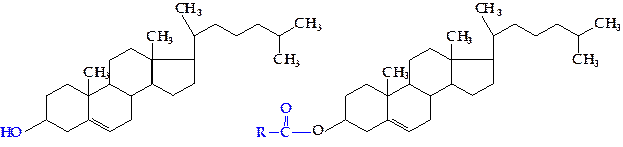
|
Холестерол служит исходным предшественником для синтеза всех стероидов, функционирующих в организме, — половых гормонов и гормонов коркового слоя надпочечников (кортикостероидов), желчных кислот, витамина D3. Кроме этого, холестерол, входящий в состав клеточных мембран, оказывает существенное влияние на состояние мембраны и таким образом участвует в регуляции ее проницаемости. Много холестерола содержится в молоке, сливочном масле, яичном желтке.
ГОРМОНЫ И ВИТАМИНЫ
Гормоны (от греч. «hormao» — «возбуждаю») — это органические молекулы, которые синтезируются в клетках эндокринных желез, транспортируются кровью к тканям и клеткам-мишеням, где выполняют физиолого-биохимические регуляторные функции. Молекулы гормонов распознаются рецепторами клеток-мишеней, влияют на синтез ферментов в клетке de novo или изменяют активность ферментов, уже имеющихся в клетке.
Эволюционно сформировались четыре системы регуляции, выполняющие функции координации тканевых ответов на изменения среды, а также осуществляющие взаимосвязь между клетками, тканями и органами.
1. Центральная и периферическая нервная система, осуществляющая проведение быстрого сигнала посредством нервных импульсов и нейромедиаторов.
2. Эндокринная система, использующая в качестве мобильных химических посредников переносящих сигналы - гормоны. Скорость сигналов относительно низка и ограничена скоростью диффузии гормонов с кровью.
3. Паракринная и аутокринная системы, осуществляющие регуляцию через различные соединения, которые секретируются в межклеточное пространство и взаимодействуют с рецепторами клеток (например, эйкозаноиды, гормоны ЖКТ, гистамин).
4. Иммунная система, действующая через специфические белки (антитела, Т-рецепторы, цитокины).
Синтез и секреция гормонов стимулируются внешними и внутренними сигналами, поступающими в ЦНС. Эти сигналы по нервным связям поступают в гипоталамус, где стимулируют синтез пептидных гормонов (рилизинг-факторов) — либеринов и статинов (рисунок 2.26).

«+» (сплошные стрелки) и «-» (пунктирные стрелки) – стимуляция и подавление
синтеза и секреции гормонов
Рисунок 2.33 - Взаимосвязь регуляторных систем организма
Либерины и статины транспортируются в переднюю долю гипофиза, где стимулируют или тормозят синтез тропных гормонов. Тропные гормоны гипофиза стимулируют синтез и секрецию гормонов периферических эндокринных желез, которые поступают в общий кровоток. Изменение концентрации метаболитов в клетках-мишенях подавляет синтез гормонов по механизму отрицательной обратной связи, действуя либо на эндокринные железы, либо на гипоталамус. Синтез и секреция тропных гормонов подавляется гормонами периферических желез.
Имеются несколько вариантов классификации гормонов, которые наглядно изображены на рисунке 2.27. Однако наиболее приемлимыми среди них являются классификация гормонов по химическому строению (белково-пептидные, стероидные гормоны и т.д.) и по месту их синтеза (гормоны гипоталамуса, гипофиза и т.д.).
В соответствии с предложенной классификацией различают также три типа сигнальных молекул:
1) нейромедиаторы (нейротрансмиттеры), представляющие собой органические сигнальные молекулы, действующие в пределах того синапса, в который они освобождаются проводником (например: ацетилхолин, норадреналин, g-аминомасляная кислота);
2) истинные (классические) гормоны, обладающие дистантным действием и регулирующие обмен веществ и физиологические функции в организме (рост клеток и тканей, их дифференцировку, кровяное давление, работу репродуктивной системы, адаптационные механизмы и др.);
3) тканевые гормоны (гормоны местного действия), которые вырабатываются, связываются с рецепторами и инактивируются в пределах ограниченного участка ткани или органа (например: простаглаидины, лейкотриены и др.).
Механизмы передачи гормонального сигнала. Несмотря на большое количество гормонов, обладающих разнообразными функциями и имеющих различные структуры, механизмы их действия на клетки-мишени в значительной степень унифицированы. Выделяют три основных механизма действия гормонов: мембранный (локальный), мембранно-внутриклеточный (или опосредованный) и цитозольный (прямой).
Мембранный механизм. Данный тип действия гормонов, заключающийся в избирательном изменении проницаемости плазматической мембраны, реализуется в месте связывания гормона с мембраной. По механизму действия гормон в данном случае выступает как аллостерический эффектор транспортных систем мембраны. Так, например, обеспечивается трансмембранный перенос глюкозы под действием инсулина, аминокислот и некоторых ионов.
Мембранно-внутриклеточный механизм. Основные циклы первого этапа передачи гормонального сигнала протекают на плазматической мембране, далее перемещаясь в цитозоль клетки. Они связаны с узнаванием и трансформацией гормонального сигнала и осуществляются при помощи сложной надмолекулярной системы в несколько этапов.

Рисунок 2.27 - Классификация гормонов
Дата добавления: 2017-08-01; просмотров: 3427;