Исходный материал карельской березы
Формовое разнообразие. Изучение карельской березы в нашей стране и за рубежом показало ярко выраженный полиморфизм ее по морфологическим признакам и текстуре древесины. Карельская береза растет деревом второй или третьей величины, кустарником с белой или темной корой, гладковатой или трещиноватой, с сильно выраженными или незначительными утолщениями, с волнистой, крупно- и мелкоузорчатой древесиной. Исходя из этого, был предложен ряд классификаций, в основе которых лежало выделение габитуальных форм.
Н.О. Соколов (1950) выделил три таких формы: высокоствольную, короткоствольную, кустообразную (см. рис 13.5 на вклейке). А.И. Владимиров (1976), А.П. Евдокимов (1978) добавили к этому некоторые морфологические особенности поверхности ствола: ребристую, крупнобугорчатую, шаровидноутолщенную и мелкобугорчатую. Известны классификации В.И. Ермакова (1986), М.Г. Романовского (1986). А.Я. Любавская (1966) выделила семь основных форм карельской березы — шесть с признаками, и одну — без признаков узорчатости. В ее классификации кроме габитуса был учтен характер утолщений ствола. Из узорчатых форм четыре относятся к древовидной группе и две — к кустовидной (см. рис. 13.6 на вклейке). При этом к формам, близким по габитусу, добавлен индекс «а» и «б» и отмечается характер узорчатости древесины.
1-а — высокоствольная крупноузорчатая форма (см. рис. 13.7 на вклейке). Деревья имеют слабосбежистый ствол с небольшими бугорками или желвачками, равномерно распределенными по стволу (хорошо заметны в молодом возрасте под тонкой берестой и почти незаметны к 30-40 годам, когда стволы деревьев покрываются толстой коркой). Узорчатая древесина формируется в основном в нижней части ствола и имеет относительно равномерно распределенный крупноузорчатый рисунок. Используется для получения декоративного облицовочного шпона.
I-б — высокоствольная шаровидноутолщеная неравномерно-узорчатая форма (см. рис. 13.8 на вклейке). Деревья с крупными шаровидными или бутылковидными утолщениями по стволу, отделенными друг от друга перехватами меньшего диаметра. Узорчатая древесина формируется в местах утолщений, в «перехватах» рисунок менее плотный или отсутствует. Используется в столярно-мебельном производстве и для изготовлении художественных изделий.
П-а — короткоствольная пятнисто-узорчатая форма (см. рис. 13.9 на вклейке). Деревья выделяются своеобразным, сильно сбежистым стволом, плохо очищенным от сучьев, либо главный ствол на высоте 2-3 м резко уменьшается в диаметре и по толщине равен боковым ветвям. Представляет особую ценность для изготовления высокодекоративного шпона и художественных изделий).
И-б — короткоствольная (лироствольная) плотно-узорчатая форма (см. рис. 13.10 на вклейке). Основной ствол разветвляется на два одинаковых по форме и диаметру ствола (на высоте 2-3 м). Двуствольные деревья образуются за счет отмирания осевого побега и замены его боковыми. Дихотомическое ветвление повторяется на каждом из двух стволов в ветвях второго и третьего порядков, и ствол напоминает лиру. Представляет особую ценность для производства художественных изделий, а также может быть использован в сто-лярно-мебельном производстве.
III — кустовидная мелко-узорчатая форма (см. рис. 13.11 на
вклейке). Характеризуется коротким стволом, разветвляющимся на
несколько стволов. Высота кустовидных форм 4-8 (12) м. Благодаря насыщенному рисунку она используется для производства художественных изделий.
IV — кустарниковая соединенно -узорчатая форма, (см. рис. 13.12
на вклейке). Растения отличаются отсутствием основного ствола.
Три — шесть стволиков, отходящие от основного корня, сильно
изогнуты и разветвлены. Листовая пластинка темно-зеленая, кожистая. На стволиках утолщения распределяются в виде мелких
желвачков. Высота не более 5 м. Древесина имеет великолепный
сильнонасыщенный узор и используется для изготовления уникальных поделок и сувениров. Редкая встречаемость этой формы и не
продолжительность жизни ограничивает ее использование даже для
этих целей.
V — полнодревесная безузорчатая форма. Деревья имеют слабосбежистый или сбежистый, гладкий или ребристый ствол с грубо-трещиноватой у комля берестой. Высота дерева достигает 20-25 м, диаметр — 60-80 см. Текстура отличается небольшой волнистостью и свилеватостью за счет сердцевинных повторений и редких одиночных аномалий, имеет муаровый оттенок и может быть использована в столярно-мебельном производстве, для облицовки стен, изготовления паркета и фанеры.
Даже столь подробная классификация не исчерпывает всего формового разнообразия карельской березы в природе. Кроме перечиcленных форм, часто встречаются короткоствольные формы с четко выраженным центральным стволом, а также: карликовые и гигантские кусты; карликовые деревца с необычными (круглыми и цилиндрическими) утолщениями на ветвях; ребристые короткоствольные и высокоствольные формы (в том числе винтовая спираль формы П-б) и др. Еще больший полиморфизм карельской березы наблюдается в культурах, где в благоприятных условиях выживают даже нежизнеспособные в естественных насаждениях особи. Кроме того, в онтогенезе у древовидных форм карельской березы наблюдается переход одной формы в другую. Так, с возрастом у формы I-а, имеющей на протяжении всего ствола на коре мелкие желвачки, могут образоваться наплывы, характерные для I-б формы; формы I, приостановив рост, становятся типичными формами II, а формы II — формами III. Все это затрудняет классификацию карельской березы. По мере перехода от древовидных форм к кустовидным текстура древесины меняется от крупно- и среднеузорчатой до мелкоузорчатой, цвет древесины — от светлого до коричневого. Соответственно увеличивается плотность и удельный вес древесины. Клинальная изменчивость перечисленных выше признаков затрудняет промышленную унификацию карельской березы, так как несмотря на общие особенности форм, каждое дерево уникально по своей текстуре.
При отборе плюсовых деревьев к ним предъявляются следующие требования:
• насыщенность текстуры древесины должны соответствовать 4-5 баллу (1 балл — узорчатость слабо выражена, 5 баллов — максимальная насыщенность текстуры);
• протяженность узорчатой части ствола должна быть достаточно большой (вне зависимости от высоты дерева), в том числе отрезок ствола с древесиной, пригодной для изготовления шпона;
• диаметр в верхнем отрубе (делового отрезка ствола) > 16 см (без коры);
• древесина без гнили и трещин.
Отбор плюсовых деревьев ведется внутри форм (в первую очередь учитывается декоративность древесины) и соответственно имеет особенности:
В основном ведут отбор высокоствольной I-а формы.
У деревьев I-б формы в перехватах между утолщениями древесина должна быть также с узорчатой текстурой (по типу I-а формы) либо перехватов не должно быть — утолщения покрывают всю нижнюю часть ствола.
Для П-а и П-б формы подбираются деревья с большим диаметром и большой протяженностью основного ствола (для этих форм).
У деревьев IIIи IV формы объем ствола и деловой древесины невелик, но зачастую именно у них самая красивая текстура. Поэтому в качестве плюсовых следует отбирать самые уникальные деревья (требования по протяженности узорчатой части ствола и диаметру снижены, прямизна делового отрезка ствола не учитывается).
При отборе плюсовых деревьев карельской березы наибольшую трудность представляет определение насыщенности текстуры древесины у растущего дерева. Если у формы I-б можно подсчитать количество и определить размер утолщений, то у мелкобугорчатых либо подсчитывают количество бугорков на коре (при помощи палетки 10 х 10 см), либо в период сокодвижения на коре вырезают «окошко» 2 х 2 см и подсчитывают число аномальных включений непосредственно на древесине. Последний способ (особенно при отборе плюсовых деревьев) применяют редко.
Отбор плюсовых деревьев ведется в естественных средневозрастных или приспевающих насаждениях. В искусственных насаждениях кандидаты в плюсовые деревья подбирают среди 10-15-летних растений, затем (с интервалом 5 лет) проводят постепенную выбраковку менее интересных экземпляров. Аттестацию плюсовых деревьев начинают с 25-30-летнего возраста. Если вести отбор плюсовых деревьев в более позднем возрасте, то зачастую из-за толстой бересты метод палетки не позволяет судить о наличии бугорков и соответственно насыщенности текстуры. На каждое плюсовое дерево заполняется паспорт по единой форме со следующими дополнительными показателями: форма дерева, протяженность узорчатой части ствола (шпоновая, с насыщенным узором, с узором средней насыщенности, со слабым узором); прямизна узорчатой части ствола; диаметр у корневой шейки, на высоте 1; 1,3; 2; 3 м (и далее). У деревьев с наплывами подсчитывается их количество, протяженность и описывается форма наплыва и ствол в перехватах. У форм IIуказывается высота основного ствола и количество вторичных стволов (у П-б — описание обоих стволов); у формы III— наличие и высота штамба, число вторичных стволов; у формы IV кроме высоты указывается длина стелющихся или сильно наклоненных стволиков и их количество, а также характеристика коры, листовой пластинки и др.
Генетические модели карельской березы. Потомство карельской березы расщепляется на узорчатые и безузорчатые особи. Чаще всего это соотношении равно 1:1. В 1954 г. Т. Ruden предложил рассматривать узорчатость карельской березы как моногенный доминантный признак (А). Согласно его гипотезе узорчатые особи гетерозиготны (Аа), а безузорчатые — рецессивны (аа) по данному признаку, гомозиготы по доминантному признаку (АА) детальны. При опылении карельской березы пыльцой березы повислой в потомстве должно быть расщепление 1:1, как при анализирующем скрещивании (Аа х аа = 1 Аа: 1аа), когда расщепление по фенотипу и генотипу совпадает. При самоопылении (Аа хАа = \аа:2Аа:\АА) доминантный ген узорчатости является летальным для АА. В потомстве остаются узорчатые и безузорчатые особи в соотношении 2:1. В классической генетике известны примеры такого взаимодействия генов при скрещивании платиновых и чернобурых лисиц. У крупного рогатого скота и кур по такому принципу наследуется коротконогость. Такие формы встречаются только в гетерозиготном состоянии, гомозиготные особи гибнут. Это пример более сложного наследования признаков, чем моногенное; он относится к плейотропному взаимодействию генов, когда один ген детерминирует развитие и фенотипическое проявление нескольких признаков. Таким образом, гипотеза Рудена дает объяснение ращепления потомства карельской березы при опылении ее березой повислой и самоопылении. Однако эта теория не объясняет полиморфизм карельской березы по высоте и текстуре древесины.
В 1975 г. А.Я. Любавская выдвинула гипотезу, согласно которой «карелистость» контролируется рецессивным геном по моногенному типу. Предполагается, что узорчатость наследуется по признаку неполного доминирования. При этом доминантные гомозиготы АА — это особи с безузорчатой древесиной; Аа — древовидные узорчатые формы; аа — кустовидные и кустарниковые формы. При самоопылении древовидных форм (Аа хАа = 1АА: 2Аа: 1аа) происходит расщепление по фенотипу 3:1 (3 — узорчатые, 1 — безузорчатая). При опылении древовидных форм пыльцой кустарниковых и кустовид-ных форм (Аа х аа = 3аа : 1Аа) в потомстве должны быть только узорчатые формы, что в эксперименте не наблюдается. При всех типах скрещивания обязательно выщепляется безузорчатая форма.
Наиболее сложную модель предложил М.Г. Романовский (1986). В ее основе лежит идея сцепленного наследования узорчатости и высоты. Предполагается, что узорчатость наследуется по моногенному типу («+» — узорчатые; «0» — безузорчатые), а высота контролируется полиаллельными генами (а1 а2, а3). В этом случае замедленный рост имеют генотипы: а1а1; аха2\ ахау Генотипы а2а2, а2аъ обладают средней быстротой роста; аъаъ — быстрорастущие особи.
Автор считает, что для районов распространения карельской березы характерно сцепление а,+, что дает медленнорастущие узорчатые особи, в то время как генотип а,0 — медленнорастущие безузорчатые. Предполагается, что ген а} выступает в роли супрессора (ингибитора) по отношению к сцепленному с ним гену узорчатости: в сочетании а3 узорчатость не проявляется. Гомозиготы по гену узорчатости гибнут, а различные взаимодействия других генов определяют полиморфизм карельской березы. Согласно этой модели при самоопылении узорчатого низкорослого дерева с генотипом а2+а3° четверть узорчатых форм гибнет, остаются две низкорослые узорчатые и одна высокоствольная безузорчатая (а2+а3°* а2а3° = Ша2а2 : : 2а2+а3°: \а3°а3). Структура семьи, полученной при опылении того же дерева пыльцой березы повислой имеет соотношение узорчатых и безузорчатых форм 1:1 (а2+а3° х а3°а3° = 2а2а3 : 2а3°а3°), т. е. расщепление соответствует модели, предложенной Т. Руденом.
Как видно из приведенных моделей, объяснить все особенности карельской березы довольно сложно. В.И. Ермаков (1979, 1986) опровергает моногенную теорию наследования узорчатости текстуры древесины. Он считает, что узорчатость контролируется более чем одним геном, т. е. данный признак полигенен. СП. Погиба (1988) подтвердила это, доказав, что высота карельской березы генетически обусловлена, коэффициент наследуемости Н2 — 0,97 и контролируется аддитивными суммарными генами А2= 0,77. Узорчатость сцеплена с высотой. Не исключается плейотропное взаимодествие генов и генов-модификаторов на фоне паратипической (экологической) среды и гормонального поля.
По мере накопления экспериментального материала и знаний о биологии карельской березы генетические модели будут совершенствоваться от простого к сложному и помогут объяснить и понять это явление.
Фенетический анализ популяций карельской березы. Генетический анализ у древесных видов по потомству затруднен, так как требует значительного времени для получения следующего поколения взрослых особей. Для изучения генетической структуры видов разработаны методики селекционно-генетического анализа природных популяций, базирующиеся на законе Харди — Вайн-берга, представляющем собой схему-формулу бесконечно большой идеальной популяции, где нет давления, отбора, мутаций, миграций, дрейфа генов и где осуществляется панмиксия. Частоты генотипов в такой популяции сохраняются постоянными. В природе таких популяций не существует, сложность генетических и экологических явлений делает невозможным анализ всей системы факторов. По мнению некоторых ученых (В.М. Роне, 1980), это приводит к упрощению популяционных моделей. Тем не менее для генетического анализа популяций предложены модернизированные методики, учитывающие или исключающие вышеперечисленные параметры. Одной из подходящих для генетической оценки полиморфизма лесных популяций оказалась методика Л Л. Животовского (1982), в основе которой лежит фенетический подход. Фен — это элементарный, генетически обусловленный морфологический признак. Основной признак фена — дискретность, неделимость при передаче потомству. Фены могут быть моногенными или являться результатом сочетания нескольких генов; неделимый фен может представлять собой коррелятивно сцепленную группу признаков. Они отражают генетическую структуру данной особи, а своей частотой — генетическую структуру популяции. Их использование делает популяционное исследование более точным и глубоким (А.В. Яблоков, 1987).
Карельская береза может быть подвергнута фенетическому анализу по методу Л.А. Животовского, так как выделенные по габитусу и форме ствола формы могут рассматриваться как маркеры для выделения морф (фенов) в популяции. Фенетический анализ популяций карельской березы был начат на кафедре селекции, генетики и дендрологии МГУЛ (СП. Погиба, 1983, 1989; Е.В. Казанцева, 1997) и продолжен в Белоруссии (Т.А. Барсукова, 1995). В анализ были включены только узорчатые формы. Изучаемые природные популяции включают наибольшее число морф (m) — четыре формы: I, II, III, IV. Частоты их встречаемости обозначаются р1, р2 ... рт и в сумме равны единице; N — объем выборки. Внутрипопуляционное разнообразие оценивается показателем м — средним числом морф в популяции:
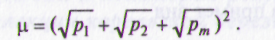
При неравномерном распределении числа морф м <т. При мономерном распределении м = 1. Ошибка Sn вычисляется по формуле
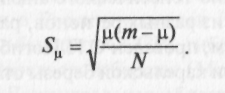
Доля редких морф определяется по формуле h = 1-м/m и оценивает структуру разнообразия популяции. Ошибка Sh вычисляется по формуле
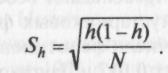
Показатель сходства популяций г вычисляется по формуле
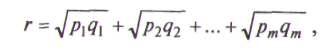
где p1, ...рт — частоты различных морф в первой популяции; q1... qm — частоты этих морф во второй популяции.
Значение показателя сходства не превышает единицы. Если сравниваемые показатели идентичны по числу морф, то r= 1. Если нет ни одной общей морфы, то r=0. Оценка значимости показателя сходства популяций проводится с помощью критерия идентичности по формуле
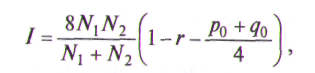
где N1 и N2 — объем выборок популяций, р0 — сумма частот морф первой выборки, не представленной во второй выборке, q0 — сумма частот морф второй выборки, не представленной в первой. Если превышает табличное значение х2 с заданным уровнем значимости и степенями свободы (m-1), то между выборками есть различие. Если rзначительно отличается от 1 (по критерию I), то его выборочная ошибка рассчитывается по формуле
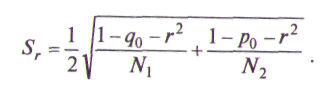
Если же каждая морфа представлена в обеих популяциях, т. е. p0=0, q0=0, формула примет вид
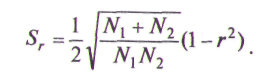
Пример популяционно-генетического анализа для семи популяций карельской березы из разных регионов, различающихся числом форм и их соотношением, проведен С.П.Погиба (1988) и Е.В. Казанцевой (1997). Популяции карельской березы отличаются друг от друга средним числом фенотипов.
Наибольшее формовое разнообразие наблюдается в Телеханском заказнике Белоруссии, среднее число фенотипов 3,66 ± 0,056, представлены все формы. Преобладают особи II формы (р2 = 0,42). Доля редких фенотипов (кустарниковых форм) незначительна — 0,085 ± 0,14. Наименьшее число фенотипов имеют латвийские популяции: Елгавская — 1,39 ± 0,017 и Даугавпилсская — 1,98 ± 0,002,но преобладают особи кустовидных форм — р3 = 0,96 и 0,60 соответственно. Редким фенотипом в обеих популяциях являются особи I высокоствольной формы.
В Заонежском заказнике Карелии преобладают особи IIIформы (р3 = 0,50), редкий фенотип — IV форма (0,175 ± 0,030); в Спасогубс-ком заказнике преобладают особи I формы (p1 = 0,70), редкий фенотип — IIформа (0,16 ± 0,035); в совхозе «Ковровский» преобладают деревья IIформы (р2 = 0,53), редкий фенотип — IIIформа (0,21 ± 0,027) и в Спас-Деменском лесхозе преобладают особи I формы (р1 = 0,60), редкий фенотип — IV форма (0,30 ± 0,026).
Различие популяций карельской березы по соотношению форм достоверно на 95 и 99% уровнях значимости (xf2 > xst2).
Данные насаждения различаются числом форм и их соотношением. Наибольшее число форм обнаружено в Телеханском заказнике, Заонежском заказнике и Спас-Деменском лесхозе, наименьшее — в Елгавском и Даугавпилсском леспромхозах (имеют только I и IIIформы).
К редким относятся IIформа в Елгавском и Даугавпилсском леспромхозах, IIформа в Спасогубском заказнике, IIIформа в совхозе «Ковровский», IV форма в Спас-Деменском межхозяйственном лесхозе, в Телеханском и Заонежском заказниках.
Выделенные популяции интересны как с эволюционно-генетичес-кой точки зрения, так и с точки зрения потребителей, поскольку являются носителями определенного генофонда и могут служить источником для селекционной работы на декоративность древесины.
13.5.3. Методы и некоторые результаты селекции карельской березы
При селекции карельской березы могут быть перспективными методы массового и индивидуального отбора; гибридизации (в том числе межвидовой); мутагенеза; полиплоидии; вегетативного размножения (в том числе микроклонирование и зеленое черенкование).
В настоящее время в России накоплен большой опыт по интродукции (14 регионов бывшего СССР), гибридизации и вегетативному размножению карельской березы (Московская область, Карелия, Костромская, Ленинградская область), заложены испытательные культуры (Московская область, Карелия), созданы заказники естественных и лучших искусственных насаждений (Карелия), отобраны плюсовые и лучшие деревья (Карелия, Московская и Костромская область). Накоплен опыт по созданию клоновых и семейственных лесосеменных плантаций (Карелия, Ленинградская обл.). В Карелии с 1934 по 1991 год практически ежегодно проводились посев (в первые годы) и посадка производственных культур, в том числе гибридного происхождения.
Ниже приводятся некоторые особенности селекции карельской березы.
Селекционная инвентаризация карельской березы. Выявление карельской березы проводится при лесоустройстве или выполнении лесохозяйственных работ. В основном, места естественного произрастания известны. Селекционная инвентаризация проводится по следующей схеме: каждое дерево карельской березы нумеруется (номер подписывается неяркой масляной краской и обязательно дублируется биркой); определяется форма по классификации А.Я. Любавс-кой, санитарное состояние дерева и необходимость в индивидуальном уходе или уходах на участке. При проведении таксационных замеров определяется: диаметр на высоте груди (D 13,м), протяженность узорчатой части ствола, высота дерева, возраст. Одновременно учитываются лучшие деревья (они описываются подробнее). Делается привязка каждого дерева или, если деревьев на участке много, составляется схема с указанием направления нумерации деревьев. Селекционная инвентаризация культур карельской березы проводится аналогично, но так как узорчатость текстуры проявляется постепенно, то ее проводят в несколько этапов. При этом постепенно вырубают безузорчатые формы, а затем, если деревья с признаками узорчатости заглушают друг друга, — и менее ценные экземпляры (со слабой текстурой и небольшим объемом ствола).
Контролируемое опыление. Для получения гибридных семян проводится опыление плюсовых деревьев по схеме диаллельного анализа, методом топкросса и др. При этом возможны различные варианты подбора родительских пар — между разными формами карельской березы или внутри форм. Контролируемое опыление проводят в урожайные годы, при обильном мужском и женском цветении и благоприятных погодных условиях в период цветения (обычно через два—три года). Если отцовские плюсовые деревья расположены далеко от материнских, целесообразно заранее провести нарезку ветвей с мужскими сережками и сделать выгонку пыльцы. Для определения жизнеспособности пыльцы ее проращивают. Опыление проводят в теплую сухую погоду. Порцию пыльцы засыпают или вдувают в изоляционный пакет из кальки, после чего пакет энергично встряхивают. Готовность женских сережек определяют по цвету — кроющие чешуйки становятся липкими и блестящими, сережка кажется не зеленой, а красноватой в пазухах кроющих чешуек. Семена собирают в конце июля (Московская область), в августе (Карелия) и хранят в холодильнике. Перед посевом, для расчетанорм высева семян, обязательно определяют всхожесть у каждого из вариантов опыления. Семена не следует хранить больше 1 года без холодильной установки.
Лесосеменные плантации. Оптимальный вариант создания лесосеменной плантации карельской березы — посадка прививок по аналогии с архивом клонов (набор клонов зависит от числа отобранных плюсовых деревьев) с расположением 5 х 8 (5 х 6) м по схеме смешения клонов (или без нее). Такая плантация одновременно является архивом клонов и служит для заготовки привойного материала.
Для выращивания посадочного материала из гибридных семян требуется незначительное по массе их количество. (Например, 100 г. семян при всхожести 60% достаточно для посева на 60 м грядки теплицы.) Поэтому на ЛСП достаточно посадить 5-10 прививок от каждого плюсового дерева. Семена получают в необходимом количестве, проводя контролируемое опыление по разным вариантам скрещивания. За прививками проводят регулярные уходы: рыхление приствольных кругов, окашивание междурядий и при необходимости, обрезку ветвей подвоя.
Стволики карельской березы повреждаются зайцами. Поэтому целесообразно на зиму укутывать их еловой лапкой. Для этого около каждой прививки вбивают три колышка высотой 1,5 м и внутрь закладывают еловую лапку. При таком укутывании прививка не сломается. После достижения растением высоты 2-2,5 м укутывание можно не проводить (если погрызы прекратились).
Испытательные культуры. Для карельской березы испытательные культуры целесообразно создавать сибсовым потомством плюсовых деревьев различных форм с применением агротехники, предложенной А.Я. Любавской: посев семян в закрытый грунт, пикировка, сортировка сеянцев по фракциям. В остальном методика создания испытательных культур общепринятая. Начиная с пятилетнего возраста, с периодичностью в 5 лет необходимо проводить раннюю диагностику. Первоначально определяют процент узорчатых и безузорчатых особей в каждой фракции каждого варианта скрещивания. Затем — распределение узорчатых особей по формам. Внутри форм проводится общая оценка узорчатых особей по 5-балльной шкале (оценка диаметра средней протяженности узорчатой части ствола и др.). В результате выявляется СКС родительских пар.
Репродукция селекционного материала, создание культур и плантаций
Вегетативное размножение. При вегетативном размножении легче добиться стабильности селектируемого признака, чем при семенном. Особое значение оно имеет при селекции методом отдаленной гибридизации и для размножения гетерозисных гибридных форм. И.О. Соколов (1950) предложил размножать стелющиеся формы карельской березы отводками. Затем опытные работы по укоренению одревесневших и зеленых черенков, в том числе с применением стимуляторов роста, были продолжены и в других регионах. При черенковании лучшие результаты получены при использовании маточников не старше 20 лет (возраст отбора кандидатов в плюсовые деревья) Кроме того, ведутся успешные опыты по клональному микроразмножению карельской березы.
Наиболее надежным методом вегетативного размножения карельской березы остается прививка. Разработку методов прививки начала А.Я. Любавская (1957). При проведении прививочных работ используют и зимние, и летние черенки. Зимние черенки с плюсовых деревьев заготавливают до начала сокодвижения. Нарезку ветвей ведут из верхней и средней части кроны, длина ветви 0,5-1 м. Летние черенки нарезаются непосредственно в день проведения прививочных работ и хранятся в воде. Различными авторами предложены и испытаны следующие способы прививки: копулировка, сердцевина на камбий, камбий на камбий, в расщеп, окулировка и др. Лучшие результаты получены при методах: на пень за кору, аблактировкой и в боковой зарез.
Прививка на пень за кору проводится в июне, черенки — зимние. Толщина подвоя у корневой шейки от 1 см и выше. Длина пенька подвоя около 15 см. Привой плотно привязывают к подвою для уменьшения вероятности облома. После того как привой приживается, подвой сажают на шип. Приживаемость прививок составляет 60% (Карелия).
Прививка аблактировкой производится летними черенками на подвой высотой 1-1,5 м. Привой длиной до 0,7 м, нижний его конец находится в емкости с водой в течение летнего периода. Осенняя приживаемость составляет около 90%, к весне следующего года она снижается до 45% (Карелия). Недостатки этого метода: ограниченное количество черенков, заготавливаемых с одного материнского дерева, сложность доставки черенков из отдаленных районов, низкая норма выработки (30-40 шт. в день).
Прививка в боковой зарез выполняется зимними черенками. Подвой — одно-двухлетние сеянцы карельской березы от контролируемого и свободного опыления высотой 0,7-1 м. Длина черенка 7-10 см. Срок прививки — июнь, обвязочный материал — полихлорвиниловая лента. Недостаток метода — частый облом привоя.
При проведении прививочных работ необходимо соблюдать следующие условия: прививки выполнять в закрытом грунте; оптималь
Альбом иллюстраций
ная температура после проведения прививочных работ и течение двух-трех недель — 18-20°С; места срезов тщательно замазывать садовым варом или пластилином; полив проводить так, чтобы вода не попадала на срезы прививаемых компонентов. Для улучшения температурного режима на период похолоданий желательно устраивать в теплице съемную «микротеплицу» над грядкой. Прижившиеся прививки карельской березы нуждаются в постоянном уходе за подвоем (регулярная обрезка побегов). Двухтрехлетние прививки можно высаживать в открытый грунт.
Агротехника выращивания семенного посадочного материала. Гибридные семена высевают в закрытый грунт с нормой расхода 1 гна 1 м2. При появлении третьего ювенильного листа у всходов целесообразно провести их пикировку в гряды с расположением 10x10 см. Как уже отмечалось, карельская береза — светолюбивая порода. При такой норме высева семян число всходов на 1 м2 может достигать 2 тыс. шт. Если не провести своевременную пикировку, подавляющее большинство их погибнет, оставшиеся будут угнетать друг друга. Отпад происходит в первую очередь за счет медленнорастущих особей, т.е. в основном будущих растений спризнаками узорчатости (особенно кустовидных и короткоствольных форм).
Пикировка позволит всем сеянцам развиваться в одинаково благоприятных условиях. Трудоемкость данной работы окупается тем, что снижается расход гибридных семян и за счет резкого уменьшения отпада сохраняются практически все растения. Без пикировки выживают в основном быстрорастущие в будущем высокоствольные формы (особенно V-безузорчатая). Осенью сеянцы выкапывают и сортируют (в каждом варианте скрещивания) на три (пять) фракций по высоте: I — самые высокие (в основном будущие 1-а, 1-6 и V безузорчатая), IIфракция — растения со средним ростом (в основном будущие короткоствольные формы), IIIфракция — растения, отставшие в росте (в основном будущие короткоствольные и кустовидные формы). Растения I и IIфракций можно использовать для посадки на плантационную или лесокультурную площадь (раздельно по фракциям). Растения IIIфракции оставляют на год на доращивание в открытом грунте.
Создание культур плантационного типа. Сеянцы, полученные из семян от контролируемого опыления по вышеприведенной агротехнике выращивания можно высаживать на лесокультурную площадь по следующим вариантам: 1) рядовой посадкой, расстояние между рядами 3,5(5) м.; 2) по 2(4) растения в посадочное место по схеме 5x5 (5x6, 5x8) м. В обоих случаях проводятся обычные лесо-хозяйственные уходы, а затем, начиная с 15-летнего возраста, селекционные рубки. Первоначально вырубаются безузорчатые особи, затем, особенно в случае групповой посадки, менее ценные растения узорчатых форм.
Раздельная посадка по фракциям (с учетом форм при вариантах скрещивания плюсовых деревьев) позволяет до некоторой степени влиять на будущий состав создаваемых культур. На участке, посаженном сеянцами от контролируемого опыления I-a x I-а форм (одна фракция по высоте), можно ожидать большего по сравнению с обычными культурами выхода высокоствольных форм, в том числе представляющей особый интерес I-а высокоствольной мелкобугристой формы, являющейся ценным сырьем для декоративного шпона. На таком участке следует сохранить и лучшие деревья 1-6, П-а и П-б форм, также дающих шпоновое сырье. По аналогии создаются участки культур из гибридных семян форм II х II (I х II, I x III, IIх III), с обязательной раздельной посадкой по фракциям.
Созданные в Карелии культуры плантационного типа (формы I х II, IIх I) в 12-летнем возрасте имели 46% узорчатых форм.
Плантации целевого назначения. Подобные плантации могут быть созданы вегетативным или семенным материалом (после испытаний на ОКС).
Плантации, созданные вегетативным материалом, позволяют по-, лучить древесину с заданными свойствами — определенной текстурой. Основное достоинство таких плантаций карельской березы — однородность древесины (по узору, цвету и др.). В то же время необходимо учитывать, что создание и выращивание таких плантаций до возраста рубки достаточно дорогостоящее мероприятие. Поэтому перспективно создавать их только для выращивания карельской березы для мебельного шпона с использованием в качестве маточников лучших плюсовых деревьев I и II форм.
Второй вариант — создание плантаций материалом, прошедшим испытание на СКС. Схемы посадки могут быть разными, в зависимости от целей селекции. Приводим один из вариантов (плюсовое дерево I-а формы х плюсовое дерево I-а формы): выращивание сеянцев; отбор I фракции, посадка по схеме 5 х 5 м по 2-4 сеянца в посадочное место; селекционные рубки в 10 лет (вырубка безузорчатых особей), в 15 лет (слабоузорчатых), в 20 лет — рубка менее ценных экземпляров узорчатых форм. При этом в каждом посадочном месте останется одно дерево карельской березы с хорошей текстурой древесины. Возможно дополнение плантаций, начиная с 10-летнего возраста.
Создание и уход за такими плантациями обходятся значительно дешевле, чем за клоновыми. Кроме того, корнесобственные имеютпреимущества по быстроте роста и устойчивости. Главный недостаток таких плантаций — разнородность по текстуре, так как каждому дереву будет присущ свой узор и другие свойства древесины.
* * *
Селекция карельской березы имеет огромное хозяйственное значение. Это одна из немногочисленных лесных пород России, имеющая мировую известность благодаря неповторимой текстуре древесины. Мебель, художественные поделки, облицовочный шпон — всегда пользуется неизменным спросом. В нашей стране разработаны и внедрены в практику эффективные методы селекции, выведения и размножения ее лучших форм. По этой технологии выращены высокопродуктивные культуры в возрасте рубки. Выращивание карельской березы может быть рентабельным в большинстве регионов нашей страны. Существующее сегодня у нас законодательство не способно защитить это ценное дерево от варварского истребления и банального воровства, что зачастую сводит на нет многолетние усилия селекционеров. Отсюда и задачи ближайшего будущего заключаются в сохранении имеющегося ценного генофонда карельской березы. В качестве первого шага ее следовало бы внести в «Красную книгу» редких и исчезающих видов и форм в нашей стране.
13.6. Селекция ольхи
Ольха используется в фанерном, мебельном, тарном и карандашном производствах, при изготовлении древесностружечных плит, паркетных досок, бездымного пороха, в гидротехнических сооружениях, для дубления кожи (ее кора содержит до 16% таннидов), для укрепления берегов рек, регулирования водотоков в поймах и др. Ценится способность ольхи фиксировать атмосферный азот. Древесные породы, высаженные в смеси с ольхой, создают насаждения более высокой продуктивности, чем чистые. В последние годы, учитывая быстроту ее роста, ольха широко используется при создании биоэнергетических плантаций и плантаций с коротким оборотом рубки в США и Западной Европе. Два вида ольхи (Alnus cordata и Alnus glutinosa) исследуются на предмет сохранения генофонда в рамках рабочей сети по благородным лиственным породам (Noble Hardwoods Network). Первым видом занимаются во Франции и Италии, a Alnus glutinosa исследуется в 16 странах Европы (Австрии, Великобритании, Франции, Германии, Италии и др.) (J. Turok et al., 1998).
Дата добавления: 2017-01-13; просмотров: 1836;