Понятие популяции. Генетическая структура популяции.
ПОПУЛЯЦИЯ (фр.population население) – это совокупность особей одного вида, свободно скрещивающихся между собой. Пространственное (территориальное) распределение особей практического каждого вида неравномерно, в результате чего они образуют ЛОКАЛЬНЫЕ ПОПУЛЯЦИИ. Такие популяции , как правило связаны между собой за счет миграции особей, перенесения пыльцы растений и семян. Изучением генетических процессов, определяющих наследственную преемственность в популяциях, составляет особую область генетики – ПОПУЛЯЦИОННАЯ ГЕНЕТИКА- это наиболее математизированная область генетики, в основе которой лежит классическая работа С.С. Четверикова «О некоторых момента эвалюционнго учения с точки зрения современной генетики» (1926). Все подходы к анализу генетических процессов, имеющих место в популяциях, основаны на исследовании так называемых панмиктических популяций- популяций состоящих из неограниченного числа особей, имеющих возможность спариваться в любых сочетаниях, независимо от их генетической природы, что определяет в целом вероятность встреч определенных генотипов. Такая популяция является как бы модельной популяцией, с которой сравнивают данные получаемые статистическими методами, при изучении конкретных популяций растений, животных, микроорганизмов и т.д.
Еще один аспект изучения популяций – харакеристика их по такому параметру, как частота определенных (изучаемых, маркерных) генов. Частота отдельных генов в популяции показывает их долю в общем генофонде, т.е. совокупности генотипов всех особей популяции. Так как частоты определенных генов в популяции при свободном спаривании определяют частоты генотипов и фенотипов, то по последним параметрам их и определяют.
Если диплоидная популяция насчитывает N особей, то генофонд состоит из 2 N гаплоидных геномов, т.е. включает в себя по 2 Nгенов каждого локуса N пар гомологичных хромосом. Исключение составляют гены, локализованные в половых хромосомах гетерогаметных особей.
Популяция является генетической единицей вида, изменения которой осуществляет эволюция вида. Как группа совместно обитающих особей одного вида, популяция выступает первой надорганизменной биологической макросистемой. У популяции приспособительные возможности значительно выше, чем у составляющих ее индивидов. Популяция как биологическая единица обладает определенными структурой и функциями.
Структура популяции характеризуется составляющими ее особями и их распределением в пространстве. Популяции свойственна определенная организация. Распределение особей по территории, соотношения групп по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают структуру популяции. Она формируется, с одной стороны, на основе общих биологических свойств вида, а с другой — под влиянием абиотических факторов среды и популяций других видов. Структура популяций имеет, следовательно, приспособительный характер. Генетическая структура популяции, определяется генофондом популяции, частотами генов и генотипов. Генофонд образован совокупностью генотипов всех особей популяции.
Генетическая структура популяции определяется изменчивостью и разнообразием генотипов, частотами вариаций отдельных генов — аллелей, а также разделением популяции на группы генетически близких особей, между которыми при скрещивании происходит постоянный обмен аллелями.
Генетическую структуру популяции мы можем описать, определив частоты генотипов в ее генофонде. Для этого нам нет необходимости обследовать всех особей этой популяции. Биологи, как правило, анализируют выборку особей из популяции. Чем больше эта выборка, тем точнее она представляет реальное соотношение частот генотипов в популяции. В качестве материала используются полевые наблюдения, данные лабораторных анализов полевых сборов или музейных экземпляров и даже архивные данные.
62. Закон Харди-Вайнберга
В 1908 г. немецкий врач В.Вайнберг и английский математик Г.Харди независимо друг от друга обосновали правило, которому подчиняются частоты распространения гомозигот и гетерозизот в панмиктических популяциях. Это правило получило название «закон Харди-Вайнберга». Основой его является утверждение, что отсутствие элементарных эволюционных процессов (мутации, миграции, отбор) частоты генов из поколения в поколение остаются неизменными. Практическое же значение уравнения в селекции – позволяет выявить генетический потенциал исходного материала (природных популяций, а также сортов и пород народной селекции), поскольку разные сорта и породы характеризуются собственными аллелофондами, которые могут быть рассчитаны с помощью закона Харди-Вайнберга. Если в исходном материале выявлена высокая частота требуемого аллеля, то можно ожидать быстрого получения желаемого результата при отборе. Если же частота требуемого аллеля низка, то нужно или искать другой исходный материал, или вводить требуемый аллель из других популяций (сортов и пород).
Уравнение Харди-Вейнберга базируется на законах Менделя. Каждая диплоидная особь в популяции возникает от слияния двух гаплоидных гамет. Следовательно частоты аллелей в популяции особей равны частотам аллелей в популяции гамет, от объединения которых эти особи возникли. Пусть доля сперматозоидов и яйцеклеток несущих аллель В равна р, а доля гамет несущих аллель b равна q. Если мы допускаем, что популяция достаточно многочисленна, и объединение гамет происходит случайно, то частоты возникших генотипов должны быть равны произведению частот гамет несущих аллели В и b. Вероятность оплодотворения яйцеклетки несущей аллель В сперматозоидом, несущим аллель В равна р хр = p2. Гомозиготы bb должны возникать с частотой q2. Слияние сперматозоида несущего В, с яйцеклеткой несущей b, происходит с частотой pq. С той же частотой происходит оплодотворение яйцеклетки, несущей В, сперматозоидом, несущим b. Тогда общая частота гетерозигот Bb равна 2pq. В общем виде мы можем записать зависимость частот генотипов от частот аллелей как
(p(B)+q(b))x(p(B)+q(b))=p2(BB) + 2pq(Bb) + q2(bb).
Эта закономерность носит статистический характер, т.е. она выявляется в том случае, если численность популяции достаточно велика. Биологический смысл этого уравнения заключается в том, что распределение частот генотипов в популяции при условии случайного скрещивания между особями однозначно определяется частотами аллелей.
Популяция, в которой соотношение генотипов соответствует уравнению Харди-Вейнберга, называется равновесной. Равновесие частот генотипов в популяции поддерживается из поколения в поколение, если не нарушаются условия выполнения статистических закономерностей , то есть если скрещивания случайны, жизнеспособность особей с разными генотипами одинакова, а также не происходит изменения частот аллелей за счет мутаций, миграций или каких-либо других факторов. Чтобы решить вопрос о том, находится ли та или иная популяция в состоянии равновесия, мы должны сравнить наблюдаемые соотношения генотипов с теми, которые должны были быть в этой популяции согласно уравнению Харди-Вейнберга.
Следствия, вытекающие из закона Харди-Вайнберга:
1. Частоты аллелей не изменяются от поколения в поколение.
Частота аллеля (А или а) в потомстве равна сумме частот генотипов гомозигот (АА или аа, соответственно) и половине частот гетерозигот (Аа), т.е. частота доминантного аллеля А = р
2 + рq = р(р+ q) = р; а = q
2 + рq = q(q + р) = q (т.к. сумма всех аллелей (гамет)равна 1, т.е. р + q = 1).
Это следствие очень важно для вычисления частоты аллели в популяции, представленной известным соотношением генотипом.
2. Частоты генотипов в панмиктической популяции не меняются в ряду поколений, так как частоты генотипов в следующем поколении, так же остаются неизменными и соответственно равными р2, 2рq, q2.
Таким образом, генотипическая структура популяции одинакова как в первом, так и в последующих поколениях при условии отсутствия давления отбора.
3. Нельзя избавиться от рецессивного аллеля в популяции. Чем меньше частота рецессивного аллеля (q), тем больше частота доминантного аллеля (р), а следовательно, увеличивается доля гетерозигот в популяции (2рq), которые с равной вероятностью образуют гаметы, содержащие рецессивный аллель(а) и доминантный аллель (А).
Поддержание равновесного состояния популяции в соответствии с формулой Харди-Вайнберга обусловлено:
- свободным скрещиванием особей внутрипопуляции;
- поддержание гетерозиготности в популяции (гетерозиготы имеют большую приспособленность, более широкий диапазон нормы реакции, а также ввиду накопления новых мутаций в гетерозиготном состоянии);
- поддережание полиморфизма в популяции. Полиморфизм популяции – это существование в ней ряда генетически различных форм, воспроизводящихся при размножении
63. Факторы динамики генетической структуры популяции.
Эволюция организмов и есть непрерывная замена одних генотипов другими. Эта смена происходит в популяции путем изменения численного соотношения качественно различающихся генотипов. Изменение соотношения генотипов составляет сущность динамики генетической структуры популяции. Генетическая изменчивость популяции складывается из мутационной и комбинативной изменчивости. Равновесие генотипов в популяции, основанное на сохранении относительных частот генов, изменяется под влиянием ряда постоянно действующих факторов, к которым относятся: 1) мутационная изменчивость, 2) действие отбора, 3) миграция, 4) изменение численности популяции, 5) избирательность спаривания и оплодотворения и ряд других факторов.
Необратимое изменение генетической структуры популяции, которое может произойти под влиянием различных факторов, называют элементарным эволюционным явлением.
Существование панмиктической популяции определяется соблюдением следующих условий:
-отсутствие мутационного процесса, влияющего на частоту аллелей
-равные шансы спаривания особей, независимо от их генотипа
-отсутствие давления отбора (естественного и искусственного)
-отсутствие давления миграции особей из других популяций
-достаточная численность особей
-отсутствие эффекта изоляции.
Соблюдение всех этих условий обеспечивает существование равновесной популяции согласно закону Харди-Вайнберга. Но ни одна популяция в современном мире не изолирована от воздействия условий ее существований, которые инициируют многочисленные процессы, приводящие к изменениям ее структуры.
Известно несколько факторов, которые могут нарушать равновесие генотипов в популяции.
• мутационный процесс
• низкая численность популяций
• избирательность скрещиваний
• миграции
• отбор
Основными процессами, имеющими наибольшее влияния, признаны мутационный процесс и отбор.
В любой популяции независимо от ее размера постоянно возникают мутации, что, естественно,сказывается на частотах определенных генов. В среднем отдельный ген мутирует с частотой 10/-5. Это небольшая величина при учете, что геном состоит из сотен генов становиться значимой. В процессе мутирования возникают как прямые, так и обратные мутации. Закрепление мутаций в популяциях а, следовательно, и изменение частот генов зависит от частоты мутирования в определенном направлении, выживаемости мутанта и, наконец, плодовитости мутантной особи. При рассмотрении растений, каждое из которых продуцирует огромное количество пыльцы и образует большое число яйцеклеток, то здесь вероятность сохранения мутантных генов в гетерозиготных генотипах значительно выше.
Отбор является одной из основных причин сдвигов частот генов в популяциях. Под отбором понимается полная или частичная элиминация из популяций особей определенного генотипа. Существует два типа отбора: естественный и искусственный. Количественным показателем скорости отбора является коэффициент отбора S. Он показывает какая часть определенных генотипов популяции не оставляет потомства, а следовательно исключается из популяции. Естественный отбор обусловлен влиянием разнообразных факторов внешней среды на отдельные организмы и на всю популяцию. Искусственный отбор осуществляет человек. Главное влияние естественного отбора на популяцию выражается в том, что он способствует повышению приспособленности организмов к различным условиям жизни. Роль естественного отбора в формировании приспособленности и эволюции видов была показана в исследованиях Дарвина. Организмы, составляющие популяцию, отличаются между собой по наследственным свойствам, и реакция их на условии среды различна. Естественный отбор, поддерживая одни генотипы и устраняя другие, приводит к разнообразию организмов в популяции. Количественной мерой интенсивности воздействия естественного отбора на популяцию служит приспособленность организмов, иногда называемая селективным или адаптивным действием отбора. Показателем приспособленности является уровень интенсивности размножения и выживания особей определенного генотипа. Приспособленность, сопровождающуюся высоким уровнем воспроизводства, принимают за единицу, а с меньшими уровнями — выражают в долях единицы. Чем выше приспособленность у особей данного генотипа, тем выше частота распространения генотипа в популяции.
В зависимости от того, какое влияние оказывает отбор на признаки, различают три типа отбора: - стабилизирующий – способствует сохранению среднего значения признака;
- дизруптивный – способствует стабилизации крайних значений признака;
- движущий, или направленный – способствует непрерывному изменению признака в определённом направлении.
Поток генов, или миграция. Поток генов представляет собой обмен генами между популяциями. Миграция изменяет частоты аллелей значительно быстрее, чем мутации. Влияние потока генов на динамику популяций тех или иных организмов зависит от скорости распространения гамет и расстояния между локальными популяциями.
Численность популяции. Концентрация генов определяется численностью популяции. Чем меньше размер популяции, тем вероятнее скрещивание между собой гетерозиготных особей, дающих в потомстве рецессивных гомозигот. И, наоборот. В малочисленной популяции отбор скорее начинает устранять вредные гены и накапливать полезные. Но при небольшой численности по каким-то случайно сложившимся обстоятельствам в ней могут исчезать(элиминироваться) аллели, обусловливающие наибольшую приспособленность, а сохраняться – дающие меньшую приспособленность. В дальнейшем при увеличении численности концентрация этих аллелей быстро возрастёт. Это явление изменения генных частот в результате действия случайных факторов называют генетическим дрейфом или генетико-автоматическими процессами. Если дрейф генов приводит к снижению приспособленности популяции, то она может вымереть.
Инбридинг. Близкородственное скрещивание неизбежно при небольшой численности популяции. Инбридинг имеет несколько следствий для популяции:
- повышение гомозиготности;
- проявление рецессивных аллелей;
- при обычно отрицательном эффекте рецессивных аллелей влечёт за собой ослабление особей (инбредная депрессия);
- повышение фенотипической изменчивости вследствие выхода в гомозиготу многих аллелей.
Изоляция. Внутривидовая изоляция популяций друг от друга означает прекращение потока генов. Если популяции остаются изолированными на протяжении ряда поколений, то они могут дивергировать по генотипической структуре, особенно если отбор в них действовал в разном направлении. И это может дать начало новым видам. Изоляцию обеспечивают географические, или территориальные, экологические (территориально-климатические, микроклиматические, сезонно-климатические) и биологические факторы. Биологические факторы в итоге основаны на генетических.
Вопрос 64.
Строение ДНК и РНК. Правило комплементарности.
ДНК И РНК нуклеиновые кислоты представляющие собой полимерные молекулы различной длины, основной структурной единицей которых является нуклеотид – сложное химическое соединение, азотистое основание, ковалентно связанное с первым атомом углерода сахара и фосфатная группа.
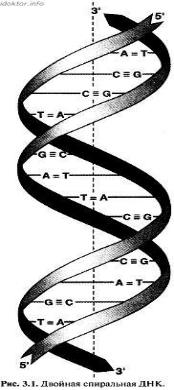
Дезоксирибонуклеи́новая кислота́ (ДНК) — макромолекула, обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. В состав ДНК входят четыре азотистых основания аденин (А), гуанин (Г) – пуриновые основания, цитозин (Ц) и тимин (Т) – пиримидиновые основания. Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т.е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК). Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м.
Первичная структура цепей ДНК — это порядок чередования дезоксирибонуклеозидмонофосфатов (dNMP) в полинуклеотидной цепи. Полинуклеотидные цепи в двухцепочечной молекуле ДНК расположены антипараллельно.
Каждая молекула ДНК упакована в отдельную хромосому. Хромосомы содержат разнообразные белки, связанные с определенными последовательностями ДНК. Все связывающиеся с ДНК эука-риотов белки можно разделить на 2 группы: гистоны и негистоновые белки. Комплекс белков с ядерной ДНК клеток называют хроматином.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула спирализована. В целом структура молекулы ДНК получила название «двойной спирали».
ДНК была открыта Иоганном Фридрихом Мишером в 1868 году. Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин, и «правил Чаргаффа», согласно которым в каждой молекуле ДНК соблюдаются строгие соотношения, связывающие между собой количество азотистых оснований разных типов[4]. Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премией по физиологии или медицине 1962 г.
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов. Первичная структура РНК — это порядок чередования рибонуклеозидмонофосфатов (NMP) в полинуклеотидной цепи. В отличии от ДНК тимин в ее составе замещен на урацил (У) , который отличается от тимина отсутствием метильной группы. Так же, как ДНК ,РНК состоит из цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков. Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК. Различают тРНК, мРНК и рРНК; все типы РНК имеют одну полинуклеотидную цепь.
Между ДНК и РНК есть три основных отличия:
ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Еще до открытия Уотсона и Крика, в 1950 г. австралийский биохимик Эдвин Чаргафф установил, что в ДНК любого организма количество адениловых нуклеотидов равно количеству тимидиловых, а количество гуаниловых нуклеотидов равно количеству цитозиловых нуклеотидов (А=Т, Г=Ц), или суммарное количество пуриновых азотистых оснований равно суммарному количеству пиримидиновых азотистых оснований (А+Г=Ц+Т). Эти закономерности получили название «правила Чаргаффа» оно и является правилом комплементарности.
Дело в том, что при образовании двойной спирали всегда напротив азотистого основания аденин в одной цепи устанавливается азотистое основание тимин в другой цепи, а напротив гуанина – цитозин, то есть цепи ДНК как бы дополняют друг друга. А эти парные нуклеотиды комплементарны друг другу (от лат. complementum – дополнение). Аденин и гуанин относятся к пуринам, а цитозин и тимин – к пиримидинам, то есть между азотистыми основаниями одной природы связи не устанавливаются. Комплементарность нуклеотидов – это химическое и геометрическое, т.е. по размерам и форме, соответствие структур их молекул друг другу.
Комплементарность позволяет зная последовательность нуклеотидов в одной спирали, выяснить порядок следования нуклеотидов на другой спирали.
Вопрос 65.
Действие гена (ДНК-РНК-полипептидная цепь).
Ген это наиболее важные в функциональном отношении части молекул ДНК. В классическом биологическом смысле ген определяли как часть хромосомы, которая обусловливает какой-нибудь один отличительный признак организма, или фенотип («фенотип» означает «внешний вид».) В дальнейшем появилось и молекулярное определение гена, впервые предложенное Джорджем Бидлом и Эдвардом Татумом в 1940 г. Они воздействовали на споры гриба Neurospora crassa рентгеновскими лучами и другими агентами, которые повреждают ДНК и таким образом вызывают мутации. Некоторые из полученных ими мутантов оказались лишенными того или иного фермента, в результате чего их метаболизм был нарушен. Это наблюдение привело авторов к заключению, что ген - это такой фрагмент генетического материала, который определяет, или кодирует, какой-то один фермент (гипотеза «один ген - один фермент»). Позже это определение гена приобрело более общий вид: «один ген - один белок», поскольку некоторые гены кодируют белки, не являющиеся ферментами. Однако теперь можно дать еще более точное биохимическое определение гена. Многие белки состоят из нескольких полипептидных цепей. В некоторых из таких белков все полипептидные цепи одинаковы; в этом случае все они могут кодироваться одним и тем же геном. Однако другие белки содержат два или большее число полипептидных цепей разных типов, различающихся по аминокислотной последовательности. Молекула гемоглобина, к примеру, состоит из полипептидных цепей двух типов-а и -р, которые различаются длиной и аминокислотной последовательностью. Сегодня мы знаем, что и-цепи кодируются двумя разными генами. Поэтому взаимосвязь между геном и белком более точно может быть выражена определением: «один ген — один полипептид».
Но не все гены экспрессируются в конечном счете с образованием полипептидных цепей. Часть генов кодирует, например, разные виды тРНК, другие гены отвечают за синтез различных рРНК. Гены, кодирующие полипептиды и РНК. называют структурными генами, ибо они определяют структуру некоего конечного продукта гена-фермента или стабильной РНК. В ДНК содержатся и другие участки или последовательности, которые выполняют исключительно регуляторную функцию. Некоторые из этих регуляторных участков представляют собой сигналы, обозначающие начало и конец структурных генов; другие принимают участие в запуске или прекращении транскрипции структурных генов. Таким образом, наряду со структурными генами хромосома содержит также регуляторные последовательности.
Современные знания о функционировании гена на молекулярном уровне могут быть обобщены в следующем виде.
1. Ген характеризуется определенным набором нуклеотидов и определенной последовательностью их в ДНК.
2. В различных генах количество нуклеотидов различно.
3. Ген непосредственно не участвует в синтезе белка, с него переписывается последовательность нуклеотидов. Переписывание лишь копирует каждый участок одной нити ДНК, соответствующий более чем одному гену.
4. Первичным продуктом гена является и-PHK с последовательностью оснований, комплементарной смысловой нити ДНК; именно этот продукт гена и является детерминирующим началом для специфического белкового синтеза. При этом само считывание гена находится также под генным контролем и контролем факторов внешней среды.
5. С шаблона гена может быть скопировано много матриц и-РНК.
Вопрос 66.
Репликация ДНК.
Репликация (позднелат. replicatio — повторение, от лат. replico — обращаюсь назад, повторяю), редупликация, ауторепродукция, аутосинтез, протекающий во всех живых клетках процесс самовоспроизведения (самокопирования) нуклеиновых кислот, генов, хромосом.
Создание модели ДНК послужило основой для понимания принципов репликации и передачи наследственной информации. ДЖ.Уотсон и Ф.Крик (1953) первыми предложили гипотезу репликации ДНК. В дальнейшем было проведено много экспериментов, результаты которых в целом подтвердили правильность модели ДНК Уотсона-Крика и их гипотезу о репликации ДНК.
Репликация ДНК — это процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты, который происходит в процессе деления клетки на матрице родительской молекулы ДНК. При этом генетический материал, зашифрованный в ДНК, удваивается и делится между дочерними клетками. Репликацию ДНК осуществляет фермент ДНК-полимераза.
В процессе репликации двойная спираль ДНК, состоящая из двух комплементарных полинуклеотидных цепей, раскручивается на отдельные цепи и одновременно начинается синтез новых полинуклеотидных цепей; при этом исходные цепи ДНК играют роль матриц. Новая цепь, синтезирующаяся на каждой из исходных цепей, идентична другой исходной цепи. Когда процесс завершается, образуются две идентичные двойные спирали, каждая из которых состоит из одной старой (исходной) и одной новой цепи. Таким образом, от одного поколения к другому передается только одна из двух цепей, составляющих исходную молекулу ДНК, – так называемый полуконсервативный механизм репликации. Репликация состоит из большого числа последовательных этапов, которые включают узнавание точки началу репликации, расплетание исходного дуплекса (спирали), удержание его цепей в изолированном друг от друга состоянии, инициацию синтеза на них новых дочерних цепей, их рост (элонгацию), закручивание цепей в спираль и терминацию (окончание) синтеза. Все эти этапы репликации, протекающие с высокой скоростью и исключительной точностью, обеспечивает комплекс, состоящий более чем из 20 ферментов и белков, – так называемая ДНК-репликазная система, или реплисома. Функциональная единица репликации – репликон, представляющий собой сегмент (участок) хромосомы или внехромосомной ДНК, ограниченный точкой начала, в которой инициируется репликация, и точкой окончания, в которой репликация останавливается. Скорость репликации контролируется на стадии инициации. Однажды начавшись, репликация продолжается до тех пор, пока весь репликон не будет дуплицирован (удвоен). Частота инициации определяется взаимодействие специальных регуляторных белков с точкой начала репликации. Бактериальные хромосомы содержат один репликон: инициации в единственной точке начала репликации ведет к репликации всего генома. В каждом клеточном цикле репликация инициируется только один раз. Плазмиды и вирусы, являющиеся автономными генетическими элементами, представляют собой отдельные репликоны, способные к многократной инициации в клетке – хозяине. Эукариотичные хромосомы (хромосомы всех организмов, за исключением бактерий и синезеленых водорослей) содержат большое число репликонов, каждый из которых также однократно инициируется за один клеточный цикл.
Начиная с точки инициации, репликация осуществляется в ограниченной зоне, перемещающейся вдоль исходной спирали ДНК. Эта активная зона репликации (т.н. репликац. вилка) может двигаться в обоих направлениях. При однонаправленной репликации вдоль ДНК движется одна репликационная вилка. При двунаправленной репликации от точки инициации в противоположных направлениях расходятся две репликационные вилки; скорости их движения могут различаться. При репликации ДНК бактерии и млекопитающих скорость роста дочерней цепи составляет соотв. 500 и 50 нуклеотидов в 1 с; у растений эта величина не превышает 20 нуклеотидов в 1 с. Движение двух вилок в противоположных направлениях создает петлю, которая имеет вид "пузыря" или "глаза". Продолжающаяся репликация расширяет "глаз" до тех пор, пока он не включит в себя весь репликон.
Вопрос 67.
Генетический код.
Генети́ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
В ДНК используется четыре азотистых основания — аденин (А), гуанин (G), цитозин (С), тимин (T), которые в русскоязычной литературе обозначаются буквами А, Г, Ц и Т. Эти буквы составляют алфавит генетического кода. В РНК используются те же нуклеотиды, за исключением нуклеотида, содержащего тимин, который заменён похожим нуклеотидом, содержащим урацил, который обозначается буквой U (У в русскоязычной литературе). В молекулах ДНК и РНК нуклеотиды выстраиваются в цепочки и, таким образом, получаются последовательности генетических букв.
После установление факта, что ДНК И РНК являются носителями наследственной информации, возник вопрос о том, как эта информация закодирована и как она пердается при построении (синтезе) белков. Данную задачу по дешифровке генетического кода полностью решили к 1966г благодаря работам 1961-1966гг. Ф Крика , С.Бреннера , Р.Холен, Х.Холена и М.Ниренберга. Ими было эксперементально показано, что каждая из 20 входящих в состав аминокислот закодирована тремя нуклеотидами, т.е. код триплетный. При синтезе(сборке) первичных цепей белка между ДНК и белком имеется проводник, который передает информацию с ДНК к белку. Этим посредником является специфическая РНК, на которую и переписывается с ДНК информация о порядке сборки аминокислот в белок. Второй особенностью кода является то, что одна аминокислота может быть закодирована не одним, а несколькими триплетами нуклеотидов и поэтому кодируется разным числом триплетов (кодонов) одним, двумя, тремя или четырмя. Эта особенность послужила основание того, что код был назван ВЫРОЖДЕННЫМ. Так же установлено, что если аминокислота кодируется двумя и более кодонами, то при синтезе белка они используются с разными частотами. Кодсчитывается в направлении от 5-крнца к 3-концу непрекращающимися триплетами, т.е. каждый кодон представлен тремя нуклеотидами, которые считываются друг за другом в строгой последовательности.
При этом весь смысл определяют два первых основания. Триплеты не перекрывают друг друга. Нуклеотид одного триплета не может входить в состав другого. Возможны любые сочетания триплетов.Разделительные знаки между триплетами отсутствуют.
Генетический код вегда описывается в символах РНК, являющейся посредником между ДНК и рибосомами, где происходит синтез белка. Старт-кодоном любого гена (началом считывания) является кодон АУГ (инициирующий кодон), а окончание считывания определяют стоп-кодоны УАА, УАГ или \уга, которые дают сигнал на окончание синтеза белка. Расстояние между стар и стоп кодонами называется открытой рамкой считывания (ORF).Надежность стоп сигнала заключается в расположении друг за другом двух или трех стоп-кадонов, первый из которых всегда УАА.
Код в целом универсален для подавляющего числа живых организмов и до недавнего времени считали, что это правило не имеет исключений, но они были обнаружены в ДНК митохондрий, и код получил название КВАЗИУНИВЕРСАЛЬНОГО.
Вопрос 68.
Типы РНК в полипептидном синтезе.
Матричная РНК
Матричная ( messenger РНК) – мРНК - посредник в передаче генетической информации от ДНК к структурам, где происходит синтез белка. Перенос информации от ДНК к мРНК называется транскрипцией и осуществляется с помощью фермента РНК-полимеразы следующим образом. В зона синтеза мРНК происходит раскручивание нити ДНК на длину 16-18 пар оснований ( два витка). Считывание идет в направлении ль 5-конца к 3-концу. Началом транскрипции является участок ДНК-промотор, а концом- терминатор, их узнают по специфическим белкам. Транскрипцию на мРНК у эукариот осуществляет фермент РНК-полимераза II, дойдя до терминатора она прекращает синтез и отделяется от матрицы. По окончании синтеза образовавшаяся мРНК переходит в цитоплазму для трансляции и прикрепляется5концрм к малой субъединице рибосомы, в которй происходит синтез белков. РНК-полимераза II локализована в нуклеосоме.
Рибосомная РНК.
Рибосома содержит 64% рибосомной РНК (рРНК) и 36% белка.Каждая из рибосом состоит из двух компонентов: малой и большой субъединиц. Их размеры были определены методом седментации и измеряются единицами S. Помолекулярной массе рРНК малые и большие субъединицы различаются.
Синтез рибосомной РНК происходит на матрице ДНК в момент образования рибосом. Этот процесс носит название транскрипции и осуществляется так же как и синтез мРНК, но в нем учавствует РНК-полимераза I, локализованная в ядрышке. В цитоплазме рибосомы, как правило располагаются группами, образуя полисомыю
Транспортная РНК.
Синтез (сборка) белка происходит в рибосомах. Для этого к месту сборки доставляются аминокислоты посредством транспортной РНК. Матрицей для синтеза тРНК являются специальные последовательности ДНК. Синтез происходит по той же схеме , но в нем учавствует РНК-полимераза II, которая содержиться в нуклеоплазме. По своей пространственной структуретРНК весьма своеобразна и напоминает конфигурацию клеверного листа. Для каждой из 20 аминокислот существует не менее одной тРНК. По нуклеотидномц составу (около 80 нуклеотидов) тРНК различны.
Дата добавления: 2017-01-13; просмотров: 6390;