Условия выполнения закона чистоты гамет
1. ход мейоза. В результате нерасхождения хромосом в одну гамету могут попасть обе гомологичные хромосомы из пары. В этом случае гамета будет нести по паре аллелей всех генов, которые содержатся в данной паре хромосом.
23. Плейотропное действие гена.
Плейотропия - явление, заключающееся в том, что один ген оказывает влияние на несколько признаков.
Плейотропия— это действие одного гена на несколько фенотипических признаков. Продукт фактически каждого гена участвует, как правило,
в нескольких, а иногда и в очень многих процессах, образующих метаболическую сеть организма. Особенно характерна плейотропия для генов, кодирующих сигнальные белки.
Плейотропи́я (от греч. πλείων — «больше» и греч. τρέπειν — «поворачивать, превращать») — явление множественного действия гена. Выражается в способности одного гена влиять на несколько фенотипических признаков. Таким образом, новая мутация в гене может оказать влияние на некоторые или все связанные с этим геном признаки. Этот эффект может вызвать проблемы при селективном отборе, когда при отборе по одному из признаков лидирует один из аллелей гена, а при отборе по другим признакам — другой аллель этого же гена.
Плейотропное действие генов - это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена.
Плейотропное действие гена может быть первичным и вторичным. При первичной плейотропии ген проявляет свой множественный эффект
При вторичной плейотропии есть один первичный фенотипний проявление гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к множественным эффектам.
При плейотропии, ген, воздействуя на какой то один основной признак, может также менять, модифицировать проявление других генов, в связи с чем введено понятие о генах-модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых "основным" геном.
24. Некумулятивная полимерия (изоэпистаз).
ПОЛИМЕРИЯ
(от греч. polymereia — многосложность), один из типов взаимодействия генов, при к-ром степень развития одного и того же признака обусловлена влиянием ряда т. н. полимерных генов (проявляющихся сходным образом). П. открыта в 1909 Н. Г. Нильсоном-Эле. П. широко распространена в природе. По типу П. наследуются ваясные хозяйственно полезные признаки: высота растений, длина вегетац. периода, количество белка в зерне, содержание витаминов в плодах, скорость протекания биохцмпч. реакций, скорость роста и масса животных, яйценоскость кур и т. д. Условно различают некумулятивную и кумулятивную П. Некумулятивная П. характеризуется тем, что для полной выраженности признака достаточно доминантного аллеля одного из полимерных генов (олнгогена).
A=B
2 гена действуют тождественно
Расщепление в F2 15:1
9 А-В- 1аавв
3 А-вв
3 ааВ-
25. Кумулятивная полимерия (полуэпистаз).
При кумулятивной П. степень выраженности признака зависит от числа доминантных аллелей как одного и того же, так и разных полимерных генов. Количеств, признаки наследуются по типу кумулятивной П. Гены действуют одинаково, они тождественны, но оба в доминантном состоянии дают суммирование своего проявления.
A=B
A+B
Расщепление 9:6:1
9А-В- 3А-вв 1аавв
3ааВ-
В основе П. на бпохимич. уровне может лежать существование неск. независимых путей биосинтеза, влияющих на развитие признака.
26. Комплементарное взаимодействие генов (коэпистаз).
A=/=B
A+B=C
Расщепление 9:3:3:1
9А-В- 3А-вв 3ааВ- 1аавв
КОМПЛЕМЕНТАРНОСТЬ
В том случае, когда признак появляется только при сочетании двух
доминантных аллелей разных генов (например, А и В) их взаимодей-
ствие называют комплементарностью, а сами гены комплементарными
(дополняющими друг друга). При этом каждый из взаимодействующих
неаллельных генов в отсутствии другого не обеспечивает формирова-
ния признака.
Комплементарное взаимодействие двух генов можно обозначить в
виде формулы:
Аа Вв
Примеры комплементарного взаимодействия генов приведены в
табл. 12.
Таблица 12
Расщепление признаков при комплементарном взаимодействии генов
Расщепле-
ние в F2
Примеры
9 : 7
Появление у душистого горошка потомства, имеющего цветки пурпур-
ной окраски, при скрещивании родительских форм с белыми цветками.
Появление у шелкопряда потомства, имеющего темно-коричневых ли-
чинок, при скрещивании неокрашенных родительских форм.
Образование цианида у растений клевера при скрещивании форм, не
образующих это соединение.
9 : 6 : 1 Появление у тыквы потомства с дисковидной формой плода при
скрещивании родителей, имеющих сферическую форму плода.
9 : 3 : 4
Появление у мышей окраски шерсти типа агути при скрещивании
родительских форм черного и белого цвета.
Появление у льна потомства с голубой окраской цветков при скре-
щивании родительских форм с неокрашенными цветками.
Появление у лука потомства с окрашенной в красный цвет лукови-
цами при скрещивании родительских форм, имеющих луковицы бе-
лого и желтого цвета.
9 : 3 : 3 : 1
Появление у дрозофилы потомства с темно-красными глазами (дикого
типа) при скрещивании родительских форм с ярко-красными (scarlet) и
коричневыми (brown) глазами.
Появление у кур потомства, имеющего ореховидный гребень при скре-
щивании родительских форм с розовидным и гороховидным гребнями.
Примечание . Жирным шрифтом выделен класс, признак которого обу-
словлен комплементарным взаимодействием генов.
27. Доминантный эпистаз.
Эпистаз
— вид взаимодействия неаллельных генов, при котором один из генов полностью подавляет действие другого гена. Ген, подавляющий действие другого гена, может называться ген-супрессор, ингибитор, эпистатичный ген. Подавляемый ген называется гипостатичным. Эпистаз может быть как доминантным, так и рецессивным. Рассмотрим пример доминантного эпистаза. У тыквы доминантный ген Y вызывает появление желтой окраски плодов, а его рецессив у — зеленой. Кроме того, имеется доминантный ген W, подавляющий проявление любой окраски, в то время как его рецессив w не мешает окраске проявляться. Поэтому растения, имеющие в своем генотипе хотя бы один доминантный ген W, будут образовывать белые плоды независимо от того, какие аллели гена Y будут присутствовать в их генотипах. Например, у растений с генотипами YYWW, YyWw, yyWw плоды будут белые.
А>B (A подавляет B)
a/>B (a не подавляет В)
A - ингибитор
а - не ингибитор
Расщепление 12:3:1
9А-В- 3ааВ- 1аавв
3А-вв
Или бывает расщепление 13:3
9А-В- 3ааВ-
3А-вв
1аавв
28. Простой и двойной рецессивный эпистаз.
Рецессивный эпистаз (recessive epistasis) [лат. recessus — отступление, удаление; греч. epistasis — неподвижность, остановка] — форма эпистаза, при которой рецессивный аллель эпистатического гена, находясь в гомозиготном состоянии, подавляет экспрессию другого (гипостатического) гена.
Двойной: аа>B
bb>A
Расщепление 9:7
9 A-B- 3A-bb
3aaB-
1aabb
Простой: аа>B
один рецессивный ген в гомозиготе подавляет доминантный другой
9:3:4
9А-В- 3А-вв 3ааВ-
1аавв
29. Сцепленное наследование признаков и генов. Группы сцепления.
Сцепленное наследование генов
Группы сцепления. Число генов у каждого организма, гораздо больше числа хромосом. Следовательно, в одной хромосоме расположено много генов. Как наследуются гены, расположенные в одной паре гомологичных хромосом?
Большую работу по изучению наследования неаллельных генов, расположенных в паре гомологичных хромосом, выполнили американский ученый Т. Морган и его ученики. Ученые установили, что гены, расположенные в одной хромосоме, наследуются совместно, или сцепленно. Группы генов, расположенные в одной хромосоме, называют группами сцепления. Сцепленные гены расположены в хромосоме в линейном порядке. Число групп сцепления у генетически хорошо изученных объектов равно числу отличающихся друг от друга хромосом. У человека 24 разновидности хромосом и 24 группы сцепления, у гороха 7 пар хромосом и 7 групп сцепления и т. д.
30. Линейное расположение генов. Карты хромосом.
Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу.
Одним из классических опытов Моргана на дрозофиле, доказывающим линейное расположение генов, был следующий. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтую окраску тела y, белый цвет глаз w и вильчатые крылья bi, были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1,2% мух кроссоверных, возникших от перекреста между генами у и w; 3,5% − от кроссинговера между генами w и bi и 4,7% — между у и bi.
Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между у и w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус.
Основным положениям хромосомной теории наследственности — парности аллелей, их редукции в мейозе и линейному расположению генов в хромосоме — соответствует однонитчатая модель хромосомы.
Существование кроссинговера побудило Моргана разработать в 1911-1914 гг. принцип построения генетических карт хромосом. В основу этого принципа положено представление о расположении генов по длине хромосомы в линейном порядке. За единицу расстояния между двумя генами условились принимать 1 % перекреста между ними.
Более обширное объяснение:
Генетические карты хромосом, схемы относительного расположения, сцепленных между собой наследственных факторов — генов. Г. к. х. отображают реально существующий линейный порядок размещения генов в хромосомах и важны как в теоретических исследованиях, так и при проведении селекционной работы, т.к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Г. к. х., можно по наследованию "сигнального" гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; например, ген, определяющий эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Многочисленные факты отсутствия (вопреки Менделя законам) независимого распределения признаков у гибридов второго поколения были объяснены хромосомной теорией наследственности. Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество которых, т. о., соответствует у каждого организма гаплоидному числу хромосом. Американский генетик Т. Х. Морган показал, однако, что сцепление генов, расположенных в одной хромосоме, у диплоидных организмов не абсолютное; в некоторых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответственными участками; этот процесс носит название перекреста, или кроссинговера. Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетический анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Г. к. х. выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему количеству изученных особей); единица этого расстояния — морганида — соответствует частоте кроссинговера в 1%.
Г. к. х. составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Г. к. х. можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы — около 400 генов, распределенных в 10 группах сцепления. У менее изученных объектов число обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, которые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления. При переносе генетического материала из клетки-донора в клетку-реципиент, например при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110—120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Г. к. х. бактерий, детально разработанных у ряда видов.
31. Кроссинговер, его значение. Типы кроссинговера.
Этот обмен участками гомологичных хромосом называется кроссинговером (перекрестом). Этот механизм появления новых генных комбинаций дополняет эффект независимой сортировки хромосом, происходящей в ходе мейоза. Различие состоит в том, что кроссинговер разделяет гены одной и той же хромосомы, тогда как независимая сортировка разделяет только гены разных хромосом.
| Кроссинго́вер (от англ. crossing over — пересечение) или перекрёст — процесс обмена участками гомологичных хромосом во время конъюгации в профазе I мейоза. Помимо мейотического, описан также митотический кроссинговер. Хромосома разделяется на эти участки в определённых точках, одних и тех же для одного вида, что может быть определением вида на генетическом уровне, место расположение этих точек задаётся единственным геном. Поскольку кроссинговер вносит возмущения в картину сцепленного наследования, его удалось использовать для картирования «групп сцепления» (хромосом). Возможность картирования была основана на предположении о том, что, чем чаще наблюдается кроссинговер между двумя генами, тем дальше друг от друга расположены эти гены в группе сцепления и тем чаще будут наблюдаться отклонения от сцепленного наследования В связи с тем, что перекомбинация генов, находящихся в одной хромосоме (сцепленных генов), обязана процессу кроссинговера, последний представляет собой важное событие с генетической точки зрения. Кроссинговер происходит в профазе мейоза I. При этом имеет место обмен гомологичными участками между хроматидами сестринских (гомологичных) хромосом. Различают такие типы кроссинговера как одинарный, двойной и множественный. На приводимой ниже схеме показаны два типа кроссинговера. Возможны и более сложные случаи обмена участками хроматид, но для генетических исследований перекомбинации генов используются эти два типа кроссинговера. Здесь следует отметить, что при этих типах кроссинговера происходит обмен равными участками хроматид с равным числом генов. Однако встречаются (редко) случаи, когда происходит разрыв хроматид в неидентичных точках и, как следствие, - обмен неравными их частями. Такой тип кроссинговера получил название неравный кроссинговер. Часто он приводит к удвоению определенных генных локусов или их нехватке. Описан также соматический кроссинговер, который происходит между хроматидами гомологичных хромосом на начальных стадиях митоза. |
32. Частота кроссинговера. Факторы, влияющие на частоту кроссинговера.
Частота (%) перекреста между двумя генами, расположенными в
одной хромосоме, пропорциональна расстоянию между ними.
Вероятность прохождения кроссинговера между двумя генами
тем реже, чем ближе друг к другу они расположены. По мере
увеличения расстояния между генами все более возрастает
вероятность того, что кроссинговер «разведет» их по двум
разным гомологичным хромосомам.
На частоту кроссинговера оказывают влияние такие факторы, как-то генотип особи, пол, возраст особи, наличие дополнительных хромосом, мутации хромосом, условия среды, при которых развивается организм и др.
33. Кроссоверные и некроссоверные гаметы.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
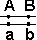
| Образуются гаметы: | 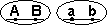
|
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
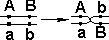
| Образуются гаметы: | 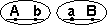
|
34. Половые хромосомы и аутосомы. Типы хромосомного определения пола.
Пол можно рассматривать как совокупность признаков и структур, обеспечивающих способ воспроизводства потомства и передачу наследственной информации. Пол чаще всего определяется в момент оплодотворения, то есть в определении пола главную роль играет кариотип зиготы. Кариотип каждого организма содержит хромосомы, одинаковые у обоих полов, — аутосомы, и хромосомы, по которым женский и мужской пол отличаются друг от друга, — половые хромосомы.
четыре типа хромосомного определения пола.
1. Женский пол — гомогаметен (ХХ), мужской — гетерогаметен (ХY) (млекопитающие, в частности, человек, дрозофила).
Генетическая схема хромосомного определения пола у человека:
| Р | ♀46, XX | × | ♂46, XY |
| Типы гамет |  23, X 23, X
| 23, X 23, Y
| |
| F | 46, XX женские особи, 50% | 46, XY мужские особи, 50% |
Генетическая схема хромосомного определения пола у дрозофилы:
| Р | ♀8, XX | × | ♂8, XY |
| Типы гамет | 4, X
| 4, X 4, Y
| |
| F | 8, XX женские особи, 50% | 8, XY мужские особи, 50% |
2. Женский пол — гомогаметен (ХХ), мужской — гетерогаметен (Х0) (прямокрылые).
Генетическая схема хромосомного определения пола у пустынной саранчи:
| Р | ♀24, XX | × | ♂23, X0 |
| Типы гамет | 12, X
| 12, X 11, 0
| |
| F | 24, XX женские особи, 50% | 23, X0 мужские особи, 50% |
3. Женский пол — гетерогаметен (ХY), мужской — гомогаметен (ХХ) (птицы, пресмыкающиеся).
Генетическая схема хромосомного определения пола у голубя:
| Р | ♀80, XY | × | ♂80, XX |
| Типы гамет | 40, X 40, Y
| 40, X
| |
| F | 80, XY женские особи, 50% | 80, XX мужские особи, 50% |
35. Особенности наследования признаков, сцепленных с полом.
Установлено, что в половых хромосомах находятся гены, отвечающие не только за развитие половых, но и за формирование неполовых признаков (свертываемость крови, цвет зубной эмали, чувствительность к красному и зеленому цвету и т.д.). Наследование неполовых признаков, гены которых локализованы в Х- или Y-хромосомах, называют наследованием, сцепленным с полом.
Изучением наследования генов, локализованных в половых хромосомах, занимался Т. Морган.
У дрозофилы красный цвет глаз доминирует над белым. Реципрокное скрещивание — два скрещивания, которые характеризуются взаимно противоположным сочетанием анализируемого признака и пола у форм, принимающих участие в этом скрещивании. Например, если в первом скрещивании самка имела доминантный признак, а самец — рецессивный, то во втором скрещивании самка должна иметь рецессивный признак, а самец — доминантный. Проводя реципрокное скрещивание, Т. Морган получил следующие результаты. При скрещивании красноглазых самок с белоглазыми самцами в первом поколении все потомство оказывалось красноглазым. Если скрестить между собой гибридов F1, то во втором поколении все самки оказываются красноглазыми, а среди самцов — половина белоглазых и половина красноглазых. Если же скрестить между собой белоглазых самок и красноглазых самцов, то в первом поколении все самки оказываются красноглазыми, а самцы белоглазыми. В F2 половина самок и самцов — красноглазые, половина — белоглазые.
Объяснить полученные результаты наблюдаемого расщепления по окраске глаз Т. Морган смог, только предположив, что ген, отвечающий за окраску глаз, локализован в Х-хромосоме (ХА — красный цвет глаз, Ха — белый цвет глаз), а Y-хромосома таких генов не содержит.
| Р | ♀XAXA красноглазые | × | ♂XaY белоглазые |
| Типы гамет |  XA XA
| Xa Y
| |
| F1 | XAXa ♀ красноглазые 50% | XАY ♂ красноглазые 50% |
| Р | ♀XAXa красноглазые | × | ♂XAY красноглазые | |
| Типы гамет | XA Xa
| XA Y
| ||
| F2 | XAXA XAXa ♀ красноглазые 50% | XАY ♂ красноглазые 25% | XaY ♂ белоглазые 25% |
| Р | ♀XaXa белоглазые | × | ♂XAY красноглазые |
| Типы гамет | Xa
| XA Y
| |
| F1 | XAXa ♀ красноглазые 50% | XaY ♂ белоглазые 50% |
| Р | ♀XAXa красноглазые | × | ♂XaY белоглазые | ||
| Типы гамет | XA Xa
| Xa Y
| |||
| F2 | XAXA ♀ красноглазые 25% | XaXa ♀ белоглазые 25% | XАY ♂ красноглазые 25% | XaY ♂ белоглазые 25% | |
У людей мужчина получает Х-хромосому от матери, Y-хромосому — от отца. Женщина получает одну Х-хромосому от матери, другую Х-хромосому от отца. Х-хромосома — средняя субметацентрическая, Y-хромосома — мелкая акроцентрическая; Х-хромосома и Y-хромосома имеют не только разные размеры, строение, но и по большей части несут разные наборы генов. В зависимости от генного состава в половых хромосомах человека можно выделить следующие участки: 1) негомологичный участокХ-хромосомы (с генами, имеющимися только в Х-хромосоме); 2) гомологичный участок Х-хромосомы и Y-хромосомы (с генами, имеющимися как в Х-хромосоме, так и в Y-хромосоме); 3) негомологичный участок Y-хромосомы (с генами, имеющимися только вY-хромосоме). В зависимости от локализации гена в свою очередь выделяют следующие типы наследования.
| Тип наследования | Локализация генов | Примеры |
| Х-сцепленный рецессивный | Негомологичный участок Х-хромосомы | Гемофилия, разные формы цветовой слепоты (протанопия, дейтеронопия), отсутствие потовых желез, некоторые формы мышечной дистрофии и пр. |
| Х-сцепленный доминантный | Негомологичный участок Х-хромосомы | Коричневый цвет зубной эмали, витамин D устойчивый рахит и пр. |
| Х-Y-сцепленный (частично сцепленный с полом) | Гомологичный участок Х- и Y-хромосом | Синдром Альпорта, общая цветовая слепота |
| Y-сцепленный | Негомологичный участок Y-хромосомы | Перепончатость пальцев ног, гипертрихоз края ушной раковины |
Большинство генов, сцепленных с Х-хромосомой, отсутствуют в Y-хромосоме, поэтому эти гены (даже рецессивные) будут проявляться фенотипически, так как они представлены в генотипе в единственном числе. Такие гены получили название гемизиготных. Х-хромосома человека содержит ряд генов, рецессивные аллели которых определяют развитие тяжелых аномалий (гемофилия, дальтонизм и пр.). Эти аномалии чаще встречаются у мужчин (так как они гемизиготны), хотя носителем генов, обусловливающих эти аномалии, чаще бывает женщина. Например, если ХА — нормальная свертываемость крови, Ха — гемофилия и если женщина является носительницей гена гемофилии, то у фенотипически здоровых родителей может родиться сын-гемофилик:
| Р | ♀XAXa норм. сверт. крови | × | ♂XAY норм. сверт. крови | |
| Типы гамет | XA Xa
| XA Y
| ||
| F2 | XAXA XАXa ♀ норм. сверт. крови 50% | XАY ♂ норм. сверт. крови 25% | XaY ♂ гемофилики 25% |
36. Пластидная наследственность. Понятия: плазмон, плазмоген.
ПЛАСТИДНАЯ НАСЛЕДСТВЕННОСТЬ, внехромосомный способ наследования пластидных признаков, осуществляемый посредством самих пластид.
В зависимости от условий оплодотворения при П. н. пластидные признаки наследуются или только по материнской линии, или от обеих родительских форм (в случае переноса пластид в зиготу и через пыльцевые трубки). О первых фактах П. н. и генетических свойствах пластид сообщили еще на заре развития генетики (1908) немецкие ботаники и генетики Э. Баур и К. Корренс изучившие наследование пестролистности у некоторых растений (герань, ночная красавица, хмель и др.). Отдельные авторы считают, что генетическими информация пластид заключена в их дезоксирибонуклеиновой кислоте (см. Нуклеиновые кислоты). Совокупность пластид клетки как структур, способных передавать наследственную информацию, названа пластидомом (О. Реннер, 1934). Из всех структурных элементов цитоплазмы растений, с которыми можно связать передачу некоторых свойств и признаков материнского организма потомству, пластиды наиболее удобны для анализа, т.к. в большинстве случаев они четко различимы в цитоплазме благодаря целому ряду морфологические особенностей. Кроме того, они способны к скачкообразным изменениям — пластидным мутациям, которые впоследствии четко воспроизводятся.
Плазмоген - носитель цитоплазматической (внеядерной) наследственности; совокупность плазмогенов образует плазмон
37. Особенности нехромосомного (цитоплазматического) наследования.
Признание за ядром главенствующей роли в передаче наследственных свойств не исключает существования внеядерной наследственности, которая связана с органоидами клетки, способными к саморепродукции. Факторы наследственности, расположенные в клетках вне хромосом, получили название плазмид. Функция плазмид, как и генов, находящихся в хромосомах, связана с ДНК. Установлено, что собственную ДНК имеют:
|
38. Цитоплазматическая мужская стерильность у растений. Использование ЦМС в селекции.
Мужская стерильность бывает при отсутствии пыльцы или неспособны ее к оплодотворению и проявляется в трех основных формах:
1. Мужская генеративные органы – тычинки – совершенно не развиваются; подобные явление наблюдается у растений некоторых видов табака;
2. Пыльники в цветках образуются, но пыльца их нежизнеспособна; эта форма стерильности чаще всего встречается у кукурузы;
3. В пыльниках образуется нормальная пыльца, но они не растрескиваются и пыльца не попадает на рыльца; это очень редкое явление наблюдается иногда у некоторых сортов томата.
Непосредственной причиной образования форм с ЦМС некоторые ученые считают нарушение синтеза белка в результате мутации в ядре, приводящей к неправильному микроспорогенезу, другие дегенерацию пыльцевых зерен связывают с нарушением снабжения питания пыльников стерильных растений.
Стерильность пыльцы, вызванная цитоплазматическими факторами, носит название цитоплазматической мужской стерильности (ЦМС). Свое название ЦМС получила потому, что этот признак наследуется по материнской линии, через цитоплазму яйцеклетки, и не передается спермиями, т. е. через мужского родителя. ЦМС возникла в ходе эволюции цветковых и проявляется обычно в популяциях растений спонтанно. Она обусловливается определенными ядерно-цитоплазматическими отношениями и выражается в дегенерации большинства пыльцевых зерен и пыльников на определенных этапах их развития. У многих видов, наиболее четко это показано для кукурузы, признак ЦМС, или стерилизующие свойства цитоплазмы, закодирован в митохондриальном геноме, а у некоторых видов растений ЦМС связывают с геномом пластид. Этот вопрос еще не совсем ясен, поскольку некоторые исследователи полагают, что ЦМС обусловлена вирусной природой.
В настоящее время это явление обнаружено у сотен видов растений и используется в связи с гетерозисом для получения гибридных семян первого поколения по принципу восстановления. Отцовская линия, как правило, несет доминантные гены и при массовой гибридизации восстанавливает фертильность в потомстве. Таким образом, высевая гибридные семена, можно получить гетерозисное потомство и фертильные семена, дающие высокий урожай растения.
№ 43, 44 МУТАЦИОННАЯ ИЗМЕНЧИВОСТЬ. ПОНЯТИЕ МУТАЦИЯ. ОСНОВНЫЕ ПОЛОЖЕНИЯ МУТАЦИОННОЙ ТЕОРИИ ДЕ-ФРИЗА.
Мутационной называется изменчивость, вызванная возникновением мутации.
Приоритет в создании теории мутаций принадлежит рус. ботанику С.И. Коржинскому, опубликовавшему в 1899г. Работу «Гетерогенезис и эволюция», и голландскому ботанику Х. де Фризу, обосновавшему в своей работе «Теория мутаций» (1901) значение мутаций (внезапных наследуемых изменений).
Мутации — это наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
Основные положения мутационной теории разработаны Г. Де Фризом в 1901—1903 гг. и сводятся к следующему:
· Мутации возникают внезапно как дискретные изменения признаков;
· Новые формы устойчивы;
· В отличие от ненаследственных изменений мутации не образуют непрерывных рядов. Они представляют собой качественные изменения;
· Мутации проявляются по-разному и могут быть как полезными, так и вредными;
· Вероятность обнаружения мутаций зависит от числа исследованных особей;
· Сходные мутации могут возникать повторно;
· Мутации не направленны (спонтанны), т. е. мутировать может любой участок хромосомы, вызывая изменения как незначительных, так и жизненно важных признаков.
Следует отметить, что любая мутация является лишь материалом для естественного отбора.
Дата добавления: 2017-01-13; просмотров: 2617;