Наука селекция и ее теоретические основы. Объекты изучения науки генетики.
Селекция (лат. selectio — выбирать) — наука о методах создания новых и улучшении существующих пород животных, сортов растений, штаммов микроорганизмов, с полезными для человека свойствами. Селекцией называют также отрасль сельского хозяйства, занимающуюся выведением новых сортов и гибридов сельскохозяйственных культур и пород животных.
В связи с развитием генетики, селекция получила новый импульс к развитию. Генная инженерия позволяет подвергать организмы целенаправленной модификации. Окончательно производится уже отбор лучших, но среди искусственно созданных генотипов.
Селекция как наука оформилась лишь в последние десятилетия. В прошлом она была больше искусством, чем наукой. Навыки, знания и конкретный опыт, нередко засекреченный, были достоянием отдельных хозяйств, переходя от поколения к поколению. Только гению Дарвина удалось обобщить весь этот огромный опыт прошлого, выдвинув идею естественного и искусственного отбора как основного фактора эволюции наряду с наследственностью и изменчивостью.
Теоретической основой селекции является генетика, так как именно знание законов генетики позволяет целенаправленно управлять появлением мутаций, предсказывать результаты скрещивания, правильно проводить отбор гибридов. В результате применения знаний по генетике удалось создать более 10000 сортов пшеницы на основе нескольких исходных диких сортов, получить новые штаммы микроорганизмов, выделяющих пищевые белки, лекарственные вещества, витамины и т. п.
К задачам современной селекции относится создание новых и улучшение уже существующих сортов растений, пород животных и штаммов микроорганизмов.
Выдающийся генетик и селекционер академик Н. И. Вавилов писал, что селекционеры должны изучать и учитывать в своей работе следующие основные факторы: исходное сортовое и видовое разнообразие растений и животных; наследственную изменчивость; роль среды в развитии и проявлении нужных селекционеру признаков; закономерности наследования при гибридизации; формы искусственного отбора, направленные на выделение и закрепление необходимых признаков.
1.Морфологическое строение и химический состав хромосом.
Хромосомы играют ведущую роль в явлениях наследственности. Они хорошо видны под световым микроскопом в момент деления клетки. Хромосомы ядра неделящейся клетки не видны, так как они деконденсированы. В то же время показано, что чем выше степень деконденсации хромосом, тем актвнее протекают метаболические процессы в самом ядре. Морфологически хромосомы растений чаще всего меют нитевидную или палочкообразную фоорму.
Большинство хромосом разделено первичной перетяжкой на 2 плеча. Под микроскопом первичная перетяжка представлена светлой зоной - центромера, которая играет основную роль в перемещении хромосом при делении ядра. Центромера занимает на каждой их хромосом строго определенное место. По положению центромеры хромосомы делят на метацентрические (приблизительно равноплечие), субметацентрические (неравноплечие) и акроцентрические (головчатые), у которых центромера сдвинута к одному из концов. У некоторых хромосом имеется и вторичная перетяжка. Она располагается у дистально конца хромосомы и отделяет небольшой ее участок, носящий название спутника. Вторичная перетяжка - ядрышковый организатор,не участвует в движении хромосом при делении ядра, поскольку в месте ее локализации происходит образование ядрышка. Концевые участки хромосомы называют теломерами. Они препятсивуют ее соединению с другими хромосомами.
Внутреннее строение хромосом. По химическому составу они на 40% состоят из ДНК и на 60% из белков, в среднем около 60% из которых приходится на гистоны. Строение метафазной хромосомы при исследовании с помощью светового микроскопа представляется следующим образом. Каждая хромосома состоит из двух хроматид, спирально закрученных и располагающихся параллельно оси хромосом хромосомы. Для прокрашивающихся в интерфазном ядре участков хромосом используют термин хромонема - красящаяся нить. Утолщения на хромонемах получили название хромомер. Особенность описанного строения хромосом зависит от уровня компактизации хроматина (комплекс ДНК с гистонами), который меняется при переходе от интерфазного состояния хромосом к метафазному.
Процесс компактизации хроматина проходит следующие уровни. Первый, нуклеосомный, определяет скрусивание ДНК по поверхности гистоновой сердцевины. Второй - объединение нескольких нуклеосом (до 10) в бусину -Т называется нуклеомерный. Третий уровень - объединение скрепками из негистоновых белков фибрилл дезоксирибонуклеопротеида в петлевой домен - хромомер. Четвертый - образование хромонем. Пятый - хромонема укладывается в виде спирали в хроматиде, уровень петлистых структур.
2.Гаплоидное и диплоидное число хромосом. Гомологичные хромосомы. Кариотип. Кариограмма.
Каждому из населяющих нашу планету видов растений и животных свойствено строго определенное число хромосом, обозначаемое число 2n (диплоидный набор). В половых клетках число хромосом в 2 раза меньше и равно n (гаплоидный набор). В соматических клетках организма каждая хромосома имеет пару, идентичную как морфологически, так и генетически (гомологичные хромосомы). Исключение из этого правила составляют половые хромосомы у гетерогаметных особей. Специфический для определенного вида по числу и структуре набор хромосом получил название кариотипа.
| Вид | Число хромосом (2n) |
| Пшеница мягкая(Triticum aestivum) | |
| Ячмень (Hordeuv vulgare) | |
| Соя (Glycine hispida) | |
| Свекла (Beta vulgaris) | |
| Верба белая (Salix alba) |
В последнее время получил распространение метод дифференциального окрашивания хромосом. При этом на каждой из хромосом прокрашиваются специфические, характерные для нее полосы (бэнды), что значительно облегчает идентификацию отдельных хромосом кариотипа. Хромосомы, определяющие пол особи, называют половыми хромосомами, а все остальные - аутосомами.
Графическое изображение Кариотипа для количественной характеристики каждой хромосомы. Один из типов К. — идиограмма — схематическая зарисовка хромосом (См. Хромосомы), расположенных в ряд по их длине (рис.). Др. тип К. — график, на котором координатами служат какие-либо значения длины хромосомы или её части и всего кариотипа (например, относительная длина хромосом) и так называемый центромерный индекс, т. е. отношение длины короткого плеча к длине всей хромосомы. Расположение каждой точки на К. отражает распределение хромосом в кариотипе. Основная задача кариограммного анализа — выявление гетерогенности (различий) внешне сходных хромосом в той или иной их группе.
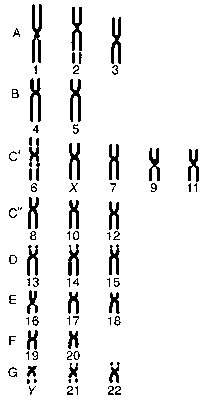
3.Деление клетки: клеточный цикл; основные периоды клеточного цикла.
Деление растительной клетки начинается с деления ядра. Деление ядра соматической клетки называется митоз. Он протекает в меристематических тканях. В результате этого деления из одной клетки образуются 2 дочерние с тем же числом хромосом, что и у родительской клетки. Между двумя клеточными делениями проходит определенный период времени, в течение которого клетка находится в состоянии видимого покоя (интерфазы). Но это только внешнее впечатление, поскольку в клетке на молекулярном уровне интенсивно протекают процессы, подготавливающие вновь образовавшуюся клетку к новому делению. Период от окончания одного митоза до окончания следующего, называют клеточным циклом. Он включает следующие периоды.
Пресинтетический (G1),в течение которого продолжается рост клеток, синтезируются специфические белки и РНК.
Синтетический (S), который характеризуется синтезом ДНК (ее количество в клетке удваивается) и гистонов. Удвоение содержания ДНК связано с репликацией хромосом. В результате в конце этого периода каждая хромосома состоит из двух хроматид.
Постсинтетический (G2), который характеризуется накоплением некоторых веществ и энергии, необходимых для протекания митоза. В этот период начинаются процессы конденсации хромосом. Перед последующим в митозе расхождением в дочерние клетки хромосомы постепенно переходят в метаболически неактивное состояние.
4.Стадии митоза. Отклонение от нормального митоза.
Подразделяется на следующие фазы.
Профазфа, на протяжении которой продолжается процесс конденсации хроматина, в результате чего хромосомы становятся видимыми под световым микроскопом, и происходит процесс разрушения ядрышка.
Метафаза, к началу которой ядерная оболочка разрушается, а хромосомы достигают максимального уровня конденсации. В это же время образуется веретено деления, состоящее из пучков протеиновых нитей, идущих от полюса к полюсу (опорные) и от полюсов к центромерам хромосом (тянущие). В результате хромосомы располагаются перпендикулярно к нитям аеретена на равном удалении от полюсов, образуя метафазную пластинку.
Анафаза ,во время которой центромеры, удерживающие сестринские хроматиды, делятся в продольном направлении и хроматиды (теперь это самостоятельные хромосомы) под действием тянущих нитей веретена начинают движение к полюсам. Деление центромер происходит синхронно. К концу анафазы в экваториальной плоскости клетки на опорных нитях веретена образуются небольшие узелки, которые в дальнейшем (по завершению телофазы) сливаются и дают начало клеточной перегородке.
Телофаза - заключительная фаза митоза. В это время начинается деконденсация хромосом, формируются ядрышки и ядерная оболочка.
Отклонения от типичного протекания митоза. Помимо митоза существуют еще 2 типа деления ядра соматических клеток: эндомитоз и амитоз.
Эндомитоз. При этом типе деления ядерная оболочка не распадается. Редупликация хромосом происходит как в митозе. В результате многократно возрастают число хромосом в ядре и размеры ядра. Эндомитоз впервые был описан в клетках тапетума шпината (Spinacia sativa).
Амитоз, или прямое деление ядра. Ядро делится на 2 части перетяжкой. Затем происходит деление цитоплазмы клетки и возникает клеточная перегородка. Амитотическое деление приводит к неравномерному распределению ДНК в дочерних клетках. Амитоз свойственен дифференцированным клеткам, таким как клетки стенок завязи, крахмалообразующие клетки клубней картофеля, клетки перисперма.
Политения. Ее можно расматривать как частный случай эндомитоза. При политении образуются гигантские хромосомы за счет многократной редупликации хроматид, но без разделения центромеры. При этом степень конденсации хроматид ниже, чем у митотических хромосом. Многочисленные хроматиды плотно прилегают друг к другу, их хромомеры образуют поперечные диски и пуффы. Впервые политенные хромосомы были обнаружены в слюнных железах личинки комара, а затем в ядрах эндосперма и антипод различных семейств растений.
5.Мейоз, фазы мейоза.
Мейоз, или редукционное деление, - особый тип деления клеток, характерный только для спорогенных тканей. При этом число хромосом в дочерних клетках уменьшается вдвое, т.е. происходит редукция числа хромосом. Мейозу предшествует интерфаза, которая аналогична таковой при митозе. В S-период интерфазы происходит редупликация хромосом, поэтому хромосомы, вступающие в процесс мейотического деления, состоят из двух хроматид. Мейоз состоит из двух ядерных делений, которые следуют одно за другим. При первом делении (мейоз ׀) происходит редукция числа хромосом, т.е. число хромосом в клетке уменьшается в 2 раза. Второе деление (мейоз ׀׀) протекает по типу митоза. Как митоз, первое и второе деления мейоза подразделяют на следующие фазы: профаза, метафаза, анафаза и телофаза. Соответственно эти фазы обозначают: метафаза ׀, метафаза ׀׀, анафаза ׀ и т.д.
Гаплоидные клетки образуются из диплоидных в результате особого клеточного деления — мейоза.
В профазе мейоза I происходит постепенная спирализация хроматина с образованием хромосом. Гомологичные хромосомы сближаются, образуя общую структуру, состоящую из двух хромосом (бивалент) и четырех хроматид (тетрада). Соприкосновение двух гомологичных хромосом по всей длине называется конъюгацией. Затем между гомологичными хромосомами появляются силы отталкивания, и хромосомы сначала разделяются в области центромер, оставаясь соединенными в области плеч, и образуют перекресты (хиазмы). Расхождение хроматид постепенно увеличивается, и перекресты смещаются к их концам. В процессе конъюгации между некоторыми хроматидами гомологичных хромосом может происходить обмен участками — кроссинговер, приводящий к перекомбинации генетического материала. К концу профазы растворяются ядерная оболочка и ядрышки, формируется ахроматиновое веретено деления. Содержание генетического материала остается прежним (2n2хр).
В метафазе мейоза I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их достигает максимума. Содержание генетического материала не изменяется (2п2хр).
В анафазе мейоза I гомологичные хромосомы, состоящие из двух хроматид, окончательно отходят друг от друга и расходятся к полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна — число хромосом уменьшается вдвое (происходит редукция). Содержание генетического материала становится 1n2хр у каждого полюса.
В телофазе происходит формирование ядер и разделение цитоплазмы — образуются две дочерние клетки. Дочерние клетки содержат гаплоидный набор хромосом, каждая хромосома — две хроматиды (1n2хр).
Интеркинез — короткий промежуток между первым и вторым мейотическими делениями. В это время не происходит репликации ДНК, и две дочерние клетки быстро вступают в мейоз II, протекающий по типу митоза.
В профазе мейоза II происходят тс же процессы, что и в профазе митоза. В метафазе хромосомы располагаются в экваториальной плоскости. Изменений содержания генетического материала не происходит (1n2хр). В анафазе мейоза II хроматиды каждой хромосомы отходят к противоположным полюсам клетки, и содержание генетического метериала у каждого полюса становится lnlxp. В телофазе образуются 4 гаплоидные клетки (lnlxp).
Таким образом, в результате мейоза из одной диплоидной материнской клетки образуются 4 клетки с гаплоидным набором хромосом. Кроме того, в профазе мейоза I происходит перекомбинация генетического материала (кроссинговер), а в анафазе I и II — случайное отхождение хромосом и хроматид к одному или другому полюсу. Эти процессы являются причиной комбинативной изменчивости.
6.Стадии профазы 1 мейоза.
Мейоз ׀ начинается с профазы ׀. Это наиболее продолжительная фаза мейоза, которая подразделяется на стадии лептотена, зиготена, пахитена, диплотена и диакинез.
На стадии лептотены в ядре появляются слабоспирализованные хромосомы. Постепенно они приобретают нитевидную форму.
Зиготена начинается с постепенного попарного соединения (конъюгации, синапсиса) по длине параллельно уложенных гомологичных хромосом. Соединенные попарно хромосомы образуют биваленты. В связи с том, что перед началом мейоза произошла редупликация хромосом, каждый бивалент состоит из четырех хроматид. Функцию синапсиса выполняет синаптонемный комплекс (СК) - белковое образование, входящее в состав бивалента и имеющее вид трехслойной ленты, располагающейся между конъюгирующими хромосомами. СК формируется постепенно по принципу застежки-молнии на протяжении всей стадии зиготены. Образование бивалентов создает предпосылки для возможности обмена гомологичными участками между гомологичными хромосомами (крассинговера), что представляет важное генетическое событие. В то же время продолжается процесс конденсации хромосом.
Пахитена - это стадия, на которой СК чформирован по всей длине хромосом (стадия стабильного синапсиса). Она характеризуется продолжающимся утолщением хромосом в результате непрерывной конденсации хроматина. На этой стадии происходит обмен гомологичными участками хроматид (кроссинговер) и, как следствие, рекомбинация сцепленных генов.
Диплотена - продолжается конденсация хромосом, но при этом начинается процесс расхождения гомологичных хромосом, которые удерживают в точках обмена участками, возникшими при кроссинговере. Они получили название хиазмы.
Диакинез - последняя стадия профазы ׀. Она характеризуется максимальной конденсацией хромосом. Исчезает ядрышко, а биваленты располагаются по периферии ядра. При этом гомологичные хромосомы удерживаются в составе бивалентов благодаря хиазам.
7.Основные различия между мейозом и митозом.
| Фаза | Митоз | Мейоз 1 деление 2 деление | |
| Интерфаза | Набор хромосом 2n Идет интенсивный синтез белков, АТФ и других органических веществ Удваиваются хромосомы, каждая оказывается состоящей из двух сестринских хроматид, скрепленных общей центромерой. | Набор хромосом 2n Наблюдаются те же процессы, что и в митозе, но более продолжительна, особенно при образовании яйцеклеток. | Набор хромосом гаплоидный (n). Синтез органических веществ отсутствует. |
| Профаза | Непродолжительна, происходит спирализация хромосом, исчезают ядерная оболочка, ядрышко, образуется веретено деления | Более длительна. В начале фазы те же процессы, что и в митозе. Кроме того, происходит конъюгация хромосом, при которой гомологичные хромосомы сближаются по всей длине и скручиваются. При этом может происходить обмен генетической информацией (перекрест хромосом) —кроссинговер. Затем хромосомы расходятся. | Короткая; те же процессы, что и в митозе, но при nхромосом. |
| Метафаза | Происходит дальнейшая спирализация хромосом, их центромеры располагаются по экватору. | Происходят процессы, аналогичные тем, что и в митозе. | Происходит то же, что и в митозе, но при nхромосом. |
| Анафаза | Центромеры, скрепляющие сестринские хроматиды, делятся, каждая из них становится новой хромосомой и отходит к противоположным полюсам. | Центромеры не делятся. К противоположным полюсам отходит одна из гомологичных хромосом, состоящая из двух хроматид, скрепленных общей центромерой. | Происходит то же, что и в митозе, но при nхромосом. |
| Телофаза | Делится цитоплазма, образуются две дочерние клетки, каждая с диплоидным набором хромосом. Исчезает веретено деления, формируются ядрышки | Длится недолго Гомологичные хромосомы попадают в разные клетки с гаплоидным набором хромосом. Цитоплазма делится не всегда. | Делится цитоплазма. После двух мейотических делений образуется 4 клетки с гаплоидным набором хромосом. |
15. Моногибридное скрещивание. Первый закон Менделя. Понятия: доминирование, рецессивность.
Моногибридное скрещивание –скрещивания родительских форм, наследственно различающихся лишь по одной паре признаков.
Первый закон Менделя - Явление единообразия гибридов первого поколения. При этом не важно, имеет ли исследователь дело с фактом полного или неполного доминирования либо со случаем кодоминирования. Во всех случаях исследователь имеет дело с единообразием особей первого гибридного поколения. Важно другое – особи (растения), вовлекаемые в скрещивания, должны быть гомозиготными.
Первый закон Менделя, получивший название закона расщепления: гибриды первого поколения F1 при дальнейшем размножении расщепляются; в их потомстве F2 снова появляются особи с рецессивными признаками, составляющие примерно четвертую часть от всего числа потомков.
Доминирование– явление преобладании признака.
Рецессивность – внешне исчезающий признак.
Закон единообразия гибридов первого поколения (первый закон Менделя) — при скрещивании двух гомозиготных организмов, относящихся к разным чистым линиям и отличающихся друг от друга по одной паре альтернативных проявлений признака, всё первое поколение гибридов (F1) окажется единообразным и будет нести проявление признака одного из родителей[3].
Этот закон также известен как «закон доминирования признаков». Его формулировка основывается на понятии чистой линии относительно исследуемого признака — на современном языке это означает гомозиготность особей по этому признаку. Понятие гомозиготности было введено позднее У. Бэтсоном в 1902 году [3].
При скрещивании чистых линий гороха с пурпурными цветками и гороха с белыми цветками Мендель заметил, что взошедшие потомки растений были все с пурпурными цветками, среди них не было ни одного белого. Мендель не раз повторял опыт, использовал другие признаки. Если он скрещивал горох с жёлтыми и зелёными семенами, у всех потомков семена были жёлтыми. Если он скрещивал горох с гладкими и морщинистыми семенами, у потомства были гладкие семена. Потомство от высоких и низких растений было высоким.
Итак, гибриды первого поколения всегда единообразны по данному признаку и приобретают признак одного из родителей. Этот признак - более сильный,доминантный (термин введён Менделем от латинского dominus), всегда подавлял другой, рецессивный[3].
Кодоминирование и неполное доминирование[править | править вики-текст]
Некоторые противоположные признаки находятся не в отношении полного доминирования (когда один всегда подавляет другой у гетерозиготных особей), а в отношении неполного доминирования. Например, при скрещивании чистых линий львиного зева с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. При неполном доминировании гетерозиготы имеют признаки, промежуточные между признаками рецессивной и доминантной гомозигот.
При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки проявляются одновременно (смешанно). Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека, где А и В — доминантные гены, а 0 — рецессивный. По этой системе генотип 00 определяет первую группу крови, АА и А0 — вторую, ВВ и В0 — третью, а АВ будет определять четвёртую группу крови. Т.о. всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвёртая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В).
Явления кодоминирования и неполного доминирования признаков слегка видоизменяет первый закон Менделя: «Гибриды первого поколения от скрещивания чистых линий особей с противоположными признаками всегда одинаковы по этому признаку: проявляют доминирующий признак, если признаки находятся в отношении доминирования, или смешанный (промежуточный) признак, если они находятся в отношении кодоминирования (неполного доминирования)».
Признаки, которые проявляются у гибридов в F1 – доминантные(преобладающие, обозначаются заглавными буквами), а альтернативные признаки - рецессивные(отсутствующие, обозначаются строчными буквами).
16. Второй закон Менделя.
Второй закон Менделя, закон независимого распределения генов – расщепление по каждой паре признаков идет независимо от других пар признаков.
Закон расщепления (второй закон Менделя) — при скрещивании двух гетерозиготных потомков первого поколения между собой во втором поколении наблюдается расщепление в определенном числовом отношении: по фенотипу 3:1, по генотипу 1:2:1.
Скрещиванием организмов двух чистых линий, различающихся по проявлениям одного изучаемого признака, за которые отвечают аллели одного гена, называется моногибридное скрещивание.
Явление, при котором скрещивание гетерозиготных особей приводит к образованию потомства, часть которого несёт доминантный признак, а часть — рецессивный, называется расщеплением. Следовательно, расщепление — это распределение доминантных и рецессивных признаков среди потомства в определённом числовом соотношении. Рецессивный признак у гибридов первого поколения не исчезает, а только подавляется и проявляется во втором гибридном поколении.
Закон чистоты гамет: в каждую гамету попадает только один аллель из пары аллелей данного гена родительской особи.
В норме гамета всегда чиста от второго гена аллельной пары. Этот факт, который во времена Менделя не мог быть твердо установлен, называют также гипотезой чистоты гамет. В дальнейшем эта гипотеза была подтверждена цитологическими наблюдениями. Из всех закономерностей наследования, установленных Менделем, данный «Закон» носит наиболее общий характер (выполняется при наиболее широком круге условий).
Гипотеза чистоты гамет. Мендель предположил, что при образовании гибридов наследственные факторы не смешиваются, а сохраняются в неизменном виде. У гибрида присутствуют оба фактора — доминантный и рецессивный, но проявление признака определяет доминантный наследственный фактор, рецессивный же подавляется. Связь между поколениями при половом размножении осуществляется через половые клетки — гаметы. Следовательно, необходимо допустить, что каждая гамета несет только один фактор из пары. Тогда при оплодотворении слияние двух гамет, каждая из которых несет рецессивный наследственный фактор, будет приводить к образованию организма с рецессивным признаком, проявляющимся фенотипически. Слияние же гамет, каждая из которых несет доминантный фактор, или же двух гамет, одна из которых содержит доминантный, а другая рецессивный фактор, будет приводить к развитию организма с доминантным признаком. Таким образом, появление во втором поколении рецессивного признака одного из родителей может быть только при двух условиях: 1) если у гибридов наследственные факторы сохраняются в неизменном виде; 2) если половые клетки содержат только один наследственный фактор из аллельной пары. Расщепление потомства при скрещивании гетерозиготных особей Мендель объяснил тем, что гаметы генетически чисты, то есть несут только один ген из аллельной пары. Гипотезу (теперь ее называют законом) чистоты гамет можно сформулировать следующим образом: при образовании половых клеток в каждую гамету попадает только один аллель из пары аллелей данного гена.
Известно, что в каждой клетке организма в большинстве случаев имеется совершенно одинаковый диплоидный набор хромосом. Две гомологичныехромосомы обычно содержат каждая по одному аллелю данного гена. Генетически «чистые» гаметы образуются следующим образом:
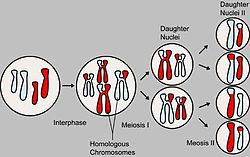
Основные этапы мейоза
На схеме показан мейоз клетки с диплоидным набором 2n=4 (две пары гомологичных хромосом). Отцовские и материнские хромосомы обозначены разным цветом.
В процессе образования гамет у гибрида гомологичные хромосомы во время I мейотического деления попадают в разные клетки. При слиянии мужских и женских гамет получается зигота с диплоидным набором хромосом. При этом половину хромосом зигота получает от отцовского организма, половину — от материнского. По данной паре хромосом (и данной паре аллелей) образуются два сорта гамет. При оплодотворении гаметы, несущие одинаковые или разные аллели, случайно встречаются друг с другом. В силу статистической вероятности при достаточно большом количестве гамет в потомстве 25 % генотипов будут гомозиготными доминантными, 50 % — гетерозиготными, 25 % — гомозиготными рецессивными, то есть устанавливается отношение 1АА:2Аа:1аа (расщепление по генотипу 1:2:1). Соответственно по фенотипу потомство второго поколения при моногибридном скрещивании распределяется в отношении 3:1 (3/4 особей с доминантным признаком, 1/4 особей с рецессивным). Таким образом, при моногибридном скрещивании цитологическая основа расщепления признаков — расхождение гомологичных хромосом и образование гаплоидных половых клеток вмейозе.
17. Понятия: Генотип и фенотип. Локус, ген, аллель. Гомозигота и гетерозигота.
Внешний вид особи (растения)- фенотип – совокупность внешних и внутренних признаков
Наследственные задатки – генотип –совокупность генов, которую организм получает от родителей
Внешний вид растения не всегда отражает наследственные задатки.
Половые гаметы несут постоянные и обособленные единицы наследственности – факторы, названные в 1909г. В. Иогансеном – генами.
Парные гены называют аллельными.
Аллели (от греч. - друг друга, взаимно) — различные формы одного и того же гена, расположенные в одинаковых участках (локусах)гомологичных хромосом и определяющие альтернативные варианты развития одного и того же признака.
Локус (лат. locus — место) в генетике означает местоположение определённого гена на генетической или цитологической карте хромосомы. Вариант последовательности ДНК в данном локусе называется аллелью.
Гомозигота —диплоидный организм или клетка, несущий идентичные аллели гена в гомологичных хромосомах.
Гетерозиготными называют диплоидные или полиплоидные ядра, клетки или многоклеточные организмы, копии генов которых в гомологичных хромосомах представлены разными аллелями. Когда говорят, что данный организм гетерозиготен (или гетерозиготен по гену X), это означает, что копии генов (или данного гена) в каждой из гомологичных хромосом несколько отличаются друг от друга.
Формы, у которых в зиготе объединяются два идентичных гена АА или аа, получили название гомозиготных, а объединяющие два разных аллеля Аа – гетерозиготных.
18. Реципрокное скрещивание.
Реципрокное скрещивание- два эксперимента по скрещиванию, характеризующиеся прямо противоположным сочетанием пола и исследуемого признака. В одном эксперименте самца, имеющего определенный доминантный признак, скрещивают с самкой, имеющей рецессивный признак. Во втором, соответственно, скрещивают самку с доминантным признаком и самца с рецессивным признаком.
Используется для определения роли пола в наследовании признака, а также позволяет определить, от какого из родителей передаются потомству цитоплазматические наследственные факторы. Для проведения реципрокного скрещивания родительские формы должны быть чистыми линиями.
РЕЦИПРОКНЫЕ СКРЕЩИВАНИЯ, обратные скрещивания, скрещивания между двумя сортами (формами) растений или породами животных, при которых каждая из родительских особей в одной комбинации скрещивания используется в качестве материнской, а в другой — в качестве отцовской формы. Как правило, в подавляющем большинстве Р. с. признаки и свойства гибрида не зависят от направления скрещивания, т. к. в обеих комбинациях Р. с. ядерный материал от родителей передается гибридам поровну. Однако цитоплазма передается только по материнской линии, и если какие-либо признаки контролируются генетически активной цитоплазмой, то она может существенно повлиять на их развитие. В таких случаях между реципрокными гибридами наблюдаются существенные различия. От выбора материнской формы часто зависит и процент завязывания гибридных семян, особенно при отдаленной гибридизации. Поэтому Р. с. применяются или с целью фиксации у реципрокных гибридов тех ценных признаков и свойств, которые генетически связаны с цитоплазмой, или когда имеются различия в репродуктивной способности гибридов в зависимости от того, в качестве материнской или в качестве отцовской берется та или иная родительская форма.
19. Анализирующее скрещивание.
Анализирующим называется скрещивание гибридной формы с родительской (например, Р2), рецессивной по изучаемому признаку:
Так как Р2 дает гаметы только одного типа, то выявление в потомстве от анализирующего скрещивания двух типов особей свидетельствует о том, что гибридная форма (F1) продуцирует гаметы двух типов, а их соотношение 1:1 – о том, что распределение доминантных и рецессивных генов в гаметах происходит с равной частотой.
Анализирующее скрещивание — скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям, то есть "анализатором". Смысл анализирующего скрещивания заключается в том, что потомки от анализирующего скрещивания обязательно несут один рецессивный аллель от "анализатора", на фоне которого должны проявиться аллели, полученные от анализируемого организма. Для анализирующего скрещивания (исключая случаи взаимодействия генов) характерно совпадение расщепления по фенотипу с расщеплением по генотипу среди потомков. Таким образом, анализирующее скрещивание позволяет определить генотип и соотношение гамет разного типа, образуемых анализируемой особью.
Мендель, проводя эксперименты по анализирующему скрещиванию растений гороха с белыми цветками (аа) и пурпурных гетерозигот (Аа), получил результат 81 к 85, что почти равно соотношению 1:1.Он определил, что в результате скрещивания и образования гетерозиготы, аллели не смешиваются друг с другом и в дальнейшем проявляются в "чистом виде". В дальнейшем Бэтсон на этой основе сформулировал правило чистоты гамет.
20. Беккросс.
Беккросс — скрещивание гибридов F1 с одной или обеими родительскими формами.
Беккросс (англ. Backcross, от back – назад и cross – скрещивание), возрастные скрещивания, скрещивания, при которых гибрид повторно (однократно или многократно) скрещивается с одним из родителей. Б. применяются для определения генотипа исследуемой форы, вычисления числа рекомбинаций или кроссинговера сцепленных генов, усиления у гибрида проявления признаков одного из родителей, а также для преодоления бесплодия гибридов 1-го поколения при отдалённой гибридизации.
Отдаленная гибридизация — межвидовая (отдаленная) гибридизация. Скрещивание особей, принадлежащих к разным видам, часто приводящее к существенному снижению жизнеспособности, частичной или полной стерильности;
21. Дигибридное скрещивание. Третий закон менделя.
Закон независимого наследования (третий закон Менделя) — при скрещивании двух особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании).
Дигибридное скрещивание- скрещивание, в которых родительские формы различаются по одной паре признаков, называются моногибридными, по двум парам – дигибридными и по многим – полигибридными.
Гены, определяющие различные признаки, наследуются независимо друг от друга. Хотя позднее было показано, что этот вывод справедлив только для генов, находящихся в разных хромосомах, закономерность, выявленная Менделем, получила название закона независимого комбинирования признаков.
Закон единообразия гибридов первого поколения (первый закон Менделя) — при скрещивании двух гомозиготных организмов, относящихся к разным чистым линиям и отличающихся друг от друга по одной паре альтернативных проявлений признака, всё первое поколение гибридов (F1) окажется единообразным и будет нести проявление признака одного из родителей[3].
Этот закон также известен как «закон доминирования признаков». Его формулировка основывается на понятии чистой линии относительно исследуемого признака — на современном языке это означает гомозиготность особей по этому признаку. Понятие гомозиготности было введено позднее У. Бэтсоном в 1902 году .
При скрещивании чистых линий гороха с пурпурными цветками и гороха с белыми цветками Мендель заметил, что взошедшие потомки растений были все с пурпурными цветками, среди них не было ни одного белого. Мендель не раз повторял опыт, использовал другие признаки. Если он скрещивал горох с жёлтыми и зелёными семенами, у всех потомков семена были жёлтыми. Если он скрещивал горох с гладкими и морщинистыми семенами, у потомства были гладкие семена. Потомство от высоких и низких растений было высоким.
Итак, гибриды первого поколения всегда единообразны по данному признаку и приобретают признак одного из родителей. Этот признак - более сильный,доминантный (термин введён Менделем от латинского dominus), всегда подавлял другой, рецессивный.
Дата добавления: 2017-01-13; просмотров: 2915;