Общая характеристика корковых движений.
Корковое движение, прирожденное или индивидуально приобретенное, в отношении координации ничем не отличается от спинномозгового двигательного рефлекса. Если при определенном слове собака становится на задние лапы и сгибает передние, то она выполняет те же координированные акты сгибания и разгибания, какими являются прирожденные спинномозговые рефлексы сгибания и разги-
бания. Или, например, если на звуковое раздражение нормальный кролик двигает ушами и поднимает голову, то и здесь мы имеем дело с такими же координированными актами, как при подобном же раздражении у кроликов, лишенных коры большого мозга. Очевидно, координация корковых и некорковых движений одна и та же и должна происходить по одному принципу. Это было доказано опытами Геринга и Шеррингтона (7), Ухтомского (43). Например, при разгибании, вызванном раздражением коры, разгибатели сокращаются, а сгибатели тормозятся (рис. 72). То же наблюдается при произвольных движениях сгибания и разгибания. Так, Бете и Каст (45) воспользовались больными, у которых была ампутирована рука, и оперировали их по методу Зауербруха — специально для ношения искусственной руки. Этот метод заключается в том, что после ампутации руки в локтевом суставе, в сухожилиях главных мышц сгибателя (m. biceps brachii) и разгибателя (m. triceps
|
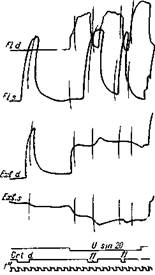
Рис.72. Выявление реципрокного торможения при корковой иннервации
Те же мышцы, что и на рис. 71. Сначала раздражается только кора правого полушария, а затем — кора в комбинации с раздражением левого чувствительного нерва n ulnarius((7. sin. 20). Последнее раздражение дает одновременно сокращение сгибателя и разгибателя на противоположной правой стороне и расслабление на разгибателе' соответствующей стороны. Под влиянием коркового раздражения правый сгибатель расслабляется одновременно с усилением сокращения правого разгибателя. В этом проявляется большая интенсивность реципрокного торможения, вызываемого из коры. Это корковое раздражение влияет тормозящим образом также на тоническое сокращение разгибателя левой стороны (Ухтомский)
brachii) делается щель, которая внутри выстилается кожей. В щель вставляются слоновые косточки, которые служат для связывания сухожилия с протезом (рис. 73—1). Таким образом, имелась возможность регистрировать сокращение антагонистов при произвольных движениях. Когда больным предлагали согнуть руку, то сгибатель сокращался, а разгибатель тормозился. Обратное наблюдалось, когда больной пытался разогнуть руку (рис. 73—II). Исследование у человека потенциалов действия антагонистических мышц при волевых движениях (Ваххольдер, 46) подтвердило, что при всяких корковых движениях сгибания и разгибания наблюдаются реципрокные отношения.
Взаимодействие между корковыми реакциями, вызываемыми непосредственно раздражением коры и разного рода спинальными и другими реакциями, совершенно такое же, как и между отдельными некорковыми реакциями. Так, например, рефлекторное сокращение мышц конечности, вызванное раздражением чувствительных нервов, ослабевает во время реципрокного коркового торможения (рис. 72). Наоборот, сами корковые реакции сильно тормозятся под влиянием таких некорковых реакций, как глотание и дефекация (Ухтомский, 43).
Весьма характерно, как это показал Введенский (47), что взаимодействие между корковыми реакциями при раздражении одноименных двигательных центров такого же рода, как при раздражении одноименных чувствительных нервов конечностей. Так, например, сокращения, вызванные слабым тетаническим раздражением двигательной зоны одного полушария, подавляются таким же раздражением одноименной зоны другого полушария. При одновременном сильном раздражении одноименных двигательных зон
|
| Рис. 73—II |
|
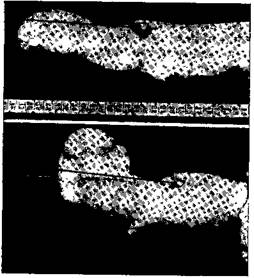
Рис. 73-1
Рис. 73—I. Ампутированная рука по способу Зауербруха
Наверху слоновая косточка проходит через сухожилие сгибателя m. biceps brachu, от нее с двух сторон идут нитки к миографу. Внизу видна такая же косточка от разгибателя m. triceps brachn. На верхнем рис. рука дана в спокойном состоянии, на нижнем — при сильном сокращении сгибателя. На верхнем рис. косточка сгибателя приходится чуть впереди косточки разгибателя, на нижнем — она отодвигается назад на несколько сантиметров. Об этом можно судить по сантиметровой шкале, помещенной между руками. Этим отодвиганием назад и осуществляется сгибание пальцев искусственной руки (Бете и Каст)
| Рис. 73—III |
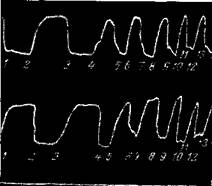
Рис. 73—II. Произвольная иннервация мышц руки, ампутированной по Зауербруху
Наверху кривая разгибателя m. triceps br., а внизу — сгибателя m. biceps br. Поочередно происходит то разгибание, то сгибание руки. Рука разгибается и сгибается по команде: «сгибатель» и «разгибатель». Все время сохраняются правильные реципрокные отношения (Бете и Каст)
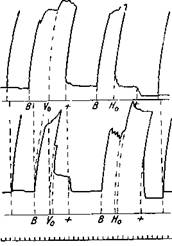
Рис. 73—III. Произвольные движения^мышц руки, ампутированной по Зауербруху
Наверху кривая разгибателя m. triceps br., внизу — сгибателя m. biceps br. При В — одновременное сокращение антагонистов; при va — изолированное сокращение разгибателя; на нем существующее сокращение усиливается, а на сгибателе оно расслабляется; при Н0 — изолированное сокращение сгибателя, разгибатель в это время расслабляется. Внизу — время в сек. (Бете)
на каждой конечности наступают ритмические сокращения, как результат смены возбуждения и торможения, подобно тому, как это позднее наблюдал Шеррингтон на спинномозговых препаратах кошки при одновременном раздражении одноименных чувствительных нервов. По-видимому, взаимодействие этих корковых реакций осуществлялось посредством координирующих аппаратов спинного мозга.
Отсюда ясно, что по внешнему проявлению нельзя судить о том, каково данное движение: корковое или некорковое. Однако, если взять не одно движение в отдельности, а все поведение животного за известный промежуток времени, то можно путем анализа определить происхождение данного движения. Так, например, произвольное корковое движение может протекать с такой последовательностью, которая не встречается среди прирожденных
рефлексов. Впрочем, существуют и такие движения, которые вызываются исключительно из коры. Таково, например, движение большого пальца у человека и обезьян и противопоставление его другим пальцам. После удаления коры это движение исчезает. Следовательно, это движение всегда является корковым.
Итак, раздражением коры мозга вызываются не только сгибателъное или разгибателъное движение конечностей, но и более или менее длительные фазы движения их. Фазные движения не имеют в коре мозга собственного представительства. Возникновение их всецело обусловливается действием вторичных проприоцептивных раздражений, ибо они исчезают после деафферентации конечностей, после перерезки соответствующих задних корешков.
Дата добавления: 2017-01-13; просмотров: 1131;