Топографическое деление коры.
Ширина всей коры, количество клеток и ширина каждого слоя клеток на фронтальном разрезе, а также клеточный состав, т. е. величина, форма, расположение клеток в каждом участке коры чрезвычайно варьируют (рис. 5). Эти вариации занимают определенные, ясно отграниченные территории. В большом мозге млекопитающих насчитывают 11 хорошо выявленных больших корковых полей. На основании гистологических и физиологических исследований каждое такое поле было разделено еще на несколько самостоятельных участков. Более значительная дифференцировка коры отмечается у высших млекопитающих — обезьян (рис. 6). Она особенно велика у человека. Поверхность коры у человека может быть разделена на 200 полей (Ц. Фохт и О. Фохт, 3).
Приводимая дифференциация в строении коры в общем совпадает с данными физиологии и клинического материала о локализации функции в коре мозга. Особенно показательны исследования неврологов Ц. Фохт и О. Фохт с электрическим раздражением коры обезьяны церкопитека. Они обнаружили полное соответствие полученных этим путем физиологических результатов с гистологически установленной архитектоникой мозга животных. На основании своих исследований эти авторы составили предположительную карту полей для человеческого мозга. Она оказалась очень сходной с той, которая была составлена О. Ферстером (4) для человеческого мозга на основании раздражения коры при операциях.

Рис. 3. Аксонные разветвления главных типов пирамидных нейронов с нисходящими аксонами(а)
/ и 5 — вставочные нейроны, их аксоны не выходят из пределов коры; 4, 7, Ю w tf — ассоциационные пирамиды, 8 — проекционный нейрон (Лоренте де Но)
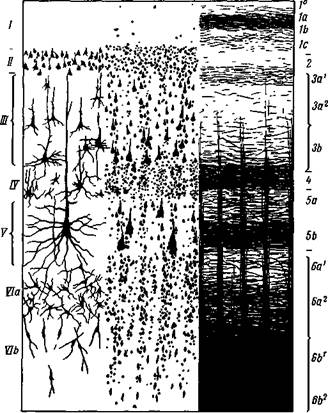
Рис. 4. Слои нервных клеток и пучков нервных волокон в коре большого мозга человека
Схематическое^изображение наиболее типичной для коры шестислойной архитектоники. С левой стороны даны клеточные слои: I — зональный или молекулярный слой; II — внешний зернистый; III — пирамидный; IV— внутренний зернистый; V — ганглионарный или внутренний пирамидный; VI — полиморфный. С правой стороны дано расположение нервных пучков в тех же слоях (по Бродману и Фохту)
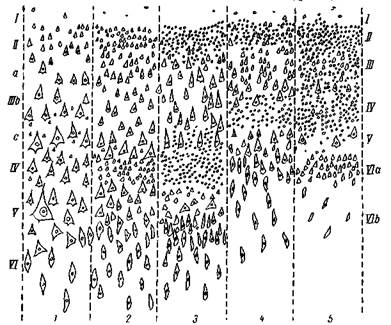
Рис. 5. Типы архитектонического строения коры больших полушарий человека
1 — тип двигательной области; 2 — фронтальный тип; з — париетальный тип; 4 — полярный тип; 5 — гранулярная, зернистая кора (Экономо)
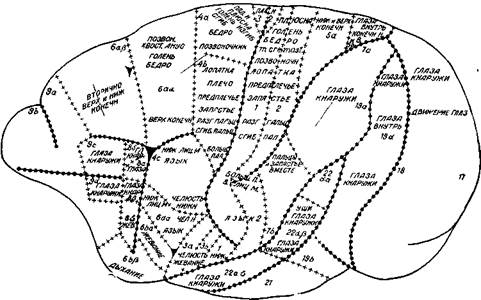
Рис. 6. Деление коры церкопитека (обезьяны) на отдельные поля на основании физиологического и гистологического исследований
Указывается, какие именно части тела реагировали на электрическое раздражение. Цифры обозначают нумерацию отдельных полей по гистологическим исследованиям (Ц. и О. Фохт)
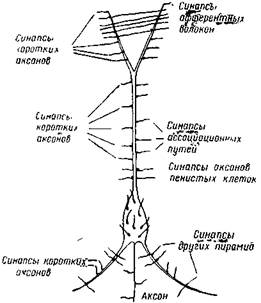
|
| Рис. 7. Пирамидный нейрон с двумя базальнымидендритами и с верхушечнымдендритом На дендритах и на теле клетки оканчивается множество синапсов от разнообразных нейронов (Лоренте Де Но) |
Микроскопическая структура коры высших позвоночных была изучена Лоренте де Но (5) на разных млекопитающих, включая и обезьяну. Подобно Р. Кахалу (6), он показал, что нейроны всех слоев коры мозга млекопитающих образуют своими сильно ветвящимися дендритами густую войлочную сеть. Многообразный нейронный состав двух наружных слоев коры мозга совсем не дает выходящих из коры аксонов. Эти аксоны клеток коротки и оканчиваются частью в том же слое, где лежат клетки, частью в соседних слоях, выше или ниже этого слоя. Иначе говоря, все нейроны первых двух слоев являются внутрикорковыми. Только в нижних четырех слоях аксоны значительного количества пирамидных клеток выходят из коры, но и здесь немало нейронов с короткими аксонами, осуществляющими связь нейронов этого слоя между собой и с вышележащими слоями. Очень характерно существование так называемых «возвратных» аксонов и коллатералей. Аксоны некоторых нейронов сначала идут вниз, а затем поворачивают обратно и заканчиваются в вышележащих слоях. Во многих случаях поворачивает обратно не весь аксон, а его коллатерали (рис. 3). Все эти аксоны и их коллатерали оканчиваются конечными или касательными синапсами в том или другом слое коры на поверхности тел клеток и их дендритов. На клетках и дендритах эти синапсы располагаются сотнями, иногда сплошным слоем (рис. 7).
Афферентные волокна, приходящие в кору из нижних отделов мозга, так называемые таламические волокна, заканчиваются преимущественно в четвертом слое, лишь немногие из них оканчиваются в третьем и пятом слоях и редко дают коллатерали к остальным слоям (рис. 1, 2). Лоренте де Но (5) находит, что эти афферентные волокна своими оканчаниями приходят в множественный синаптический контакт с находящимися здесь дендритами и телами клеток. Отсюда следует, что пирамидные нейроны нижних слоев коры могут иметь прямую связь с афферентными таламическими волокнами главным образом в своей дендритной части, распространяющейся во втором и третьем слоях. Но ввиду многочисленных аксонных связей вставочных пирамидных нейронов второго, третьего и четвертого слоев с пирамидами нижних слоев эти последние должны быть связаны с афферентными волокнами также и через эти нейроны.
Нервные круги.
Из данного краткого описания видно, что в коре большого мозга, так же как и в других отделах мозга, нервные элементы образуют замкнутые круги разной сложности. Каждый такой круг имеет свою группу афферентных и эфферентных волокон и по предположению Лоренте де Но представляет самостоятельную физиологическую единицу. В этой единице возбуждение может распространяться во всех направлениях, как от афферентного волокна к эфферентному, так и наоборот, хотя на каждом участке данной единицы импульсы возбуждения могут проводиться только в одном направлении.
По Лоренте де Но, в образовании нервных кругов принимают участие и дендриты, в особенности верхушечные. Но многочисленными физиологическими опытами доказано, что дендриты не проводят возбуждения без декремента, по закону «все или ничего». Они реагируют на нервные импульсы, приходящие через аксодендритные синапсы только локальными процессами возбуждения. Поэтому нервные круги состоят только из клеток и нейритов, т. е. аксонов.
Нервные круги образуются в коре как в вертикальном направлении, т. е. перпендикулярно к поверхности коры между нервными элементами разных слоев, так и в горизонтальном направлении, так сказать, транскортикально, между ближайшими элементами данного слоя коры. Такие связи осуществляются аксонами вставочных пирамидных нейронов. Между разными более или менее отдаленными участками коры нервные круги образуются через подкорковое белое вещество, по которому проходят аксоны ассоциационных пирамидных нейронов. Аксоны таких же нейронов связывают оба полушария через мозолистое тело. Эти пути называются каллозальными. Наконец, кора связывается с подкорковыми образованиями точно так же замкнутыми нервными кругами — кортико-таламо-кортикальными, кортико-ретикуло-кортикальными и т. д.
Нисходящие корковые пути. Нервные волокна, нисходящие из новой коры мозга, проходят сначала через белое подкорковое вещество. Часть этих волокон берет начало в гигантских пирамидных клетках Беца, которые лежат в пятом корковом слое двигательной области. Они направляются в спинной мозг и называются пирамидными путями. Эти пути проходят через ножки большого мозга, варолиев мост и продолговатый мозг. В последнем они частично перекрещиваются. Одна часть волокон переходит на другую сторону, а потом через боковые столбы оканчивается в спинном мозге. Другая часть остается на той же стороне и передними столбами вступает в спинной мозг (рис. 8). Эти пирамидные пути перерождаются, если удалить моторную кору.
Наряду с пирамидными пучками, достигающими спинного мозга без перерыва, существуют другие корковые пучки, прерывающиеся в головном мозге. Они начинаются во многих участках коры, а именно в полях 1, 2, 3, 4, 5, 6, 8 и 22, приведенных на рис. 6. Почти все ядра головного мозга получают нервные волокна из этих корковых слоев, например зрительные бугры, красные ядра, ядра четверохолмия, варолиева моста и продолговатого мозга (Терцуоло и Эди, 28). Эти пути называются экстрапирамидными (рис. 8, Б и В).
Исследованиями Монакова (7) и Ариенс Капперса (8) было установлено, что корковые центробежные волокна прерываются в спинном мозге около тех же интраспинальных и двигательных нейронов, что и коллатерали чувствительных заднекорешковых волокон. В последнее время микроэлектрофизиологические исследования подтвердили, что основная часть аксонов, идущих из головного мозга, оканчивается на вставочных нейронах промежуточной зоны и в меньшей степени — на нейронах заднего и переднего рогов. Что касается спинного мозга кошки, то для него вообще не показаны контакты пирамидных путей с мотонейронами (Кюйперс и др., 10). Но у обезьян часть волокон в области шейного утолщения оканчивается непосредственно на мотонейронах (Ниберг-Хансон и Бродал, 9). Подробные свег дения были получены по этим вопросам Уоллом (11) для поясничного отдела спинного мозга кошки. На основании экстраклеточного изучения ответов одиночных нейронов в спинном мозге им предложена схема, по которой кожные афференты заканчиваются на промежуточных нейронах, расположенных более дорзально, а аксоны бокового пучка пирамидного тракта на более вентральных и на тех, на которых заканчиваются афференты от проприорецепторов. Однако промежуточные нейроны всех плоскостей заднего рога прямо или опосредованно могут активироваться нисходящими импульсами пирамидного тракта.
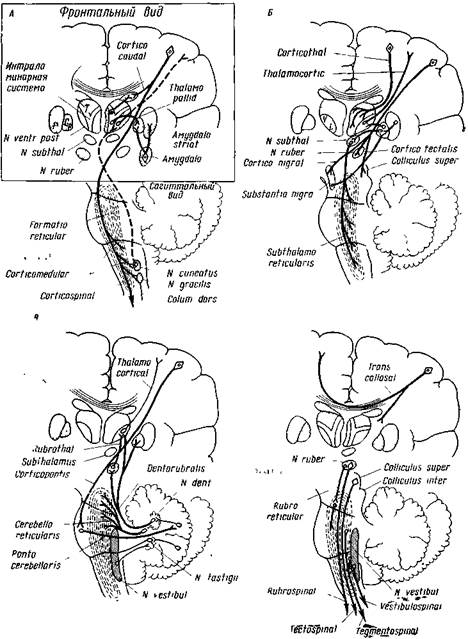
Рис. 8. Схемы кортико-субкортикальных связей
На каждом рисунке кора и подкорковые ядра даны во фронтальном, а ствол и мозжечок изображеныв сагиттальном разрезе А — первичные сенсорные пути (штрихами) и кортико-спиналыше, кортиксмедуллярные, кортико-стриарные и амигдало-стриарные пути. Б — кортико-таламические, кортикорубральные, кортико-нигральные и рубро-ретикулярные связи. В —кортико-понто-церебеллярные, церебелло-ретикулярные, церебелло-вестибулярные и церебелло-рубро-таламические пути. Г — происхождение главных субкортикальных двигательных путей к спинному мозгу: рубро-спиналъвых, тектоспинальных, тегменто-спинальных и вестибуло-спинальных(Терцуоло и Эди)
По наблюдениям Мунка (12), между рефлекторными координирующими механизмами и определенной частью двигательной сферы противоположного' полушария существует тесная связь, какой нет между той же двигательной сферой и другими рефлекторными центрами.
Однако отсюда не следует, что после разрыва какого-либо пирамидного пучка определенный двигательный орган должен потерять способность к корковым реакциям. Каждый координирующий аппарат действует не на один только двигательный орган, например, не на одну, а на несколько конечностей. Поэтому любой двигательный орган может приходить в действие не только через один наиболее прямой пирамидный путь, но и через ряд других пирамидных путей, оканчивающихся в разных координирующих аппаратах. Кроме того, известно, что двигательные органы приходят в действие также через экстрапирамидные пучки, которые прерываются в стволовой части мозга, например, в красных ядрах. Через нервные волокна, образующие нисходящие пути из данных ядер, производятся такие же движения, как и через прямые корковые пучки. Следовательно, перерезка прямых пирамидных пучков, оканчивающихся в спинном мозге, не устраняет способности к «произвольным» и вообще корковым движениям.
Прямые пирамидные пути особенно сильно развиты у человека и обезьян. Это зависит от степени развития коры больших полушарий. Так, у птиц прямые пирамидные пути развиты меньше, чем у млекопитающих. У низших позвоночных сильнее развиты непрямые пути, прерывающиеся в красных ядрах. Среди млекопитающих у человека и обезьян начинающиеся в красных ядрах монаковские нисходящие пути развиты значительно слабев, чем у собаки и кошки или у других низших млекопитающих. Если на одной стороне перерезать ножку большого мозга, а на другой — боковой столб в спинном мозге, где проходит монаковский пучок, то на стороне с перерезанным боковым столбом способность к «произвольным» движениям ослабевает больше, чем при перерезке только ножки мозга. Этим доказывается участие монаковского пучка в «произвольных» движениях. Но не он один участвует в «произвольных» движениях: после означенной двойной перерезки животное еще сохраняет способность к этим движениям. Кроме того, двигательный эффект можно вызвать путем искусственного раздражения коры. Отсюда очевидно, что из коры большого мозга двигательная реакция может быть вызвана на периферии через любой корковый путь — как через пирамидный, непосредственно оканчивающийся в спинном мозге, так и через экстрапирамидные, прерывающиеся в ядрах головного мозга.
Дата добавления: 2017-01-13; просмотров: 1534;