Основные химические канцерогены
| Группы веществ | Представители групп |
| Полициклические ароматические углеводороды | Бензопирен, диметилбензантрацен |
| Ароматические амины | 2-Ацетиламинофлуорен, К-метил-4-аминоазобензол |
| Нитрозамины | Диметилнитрозамин, диэтилнитрозамин |
| Алкилирующие агенты | Циклофосфамид, диэтилстильбэстрол |
| Природные вещества | Дактиномицин, афлатоксин B1 |
| Неорганические вещества | Хром, бериллий, асбест, свинец, кадмий |
Канцерогенным действием обладает огромное количество различных по химическому строению веществ. В печени большинство из этих веществ проканцерогены - соединения, не взаимодействующие с генетическим аппаратом клеток. После дополнительной метаболической модификации они превращаются в канцерогены, способные реагировать с молекулами нуклеиновых кислот и белков, нарушать работу регуляторных механизмов клеток и вызывать рост опухолей. Трансформация клеток под действием канцерогенов получила название химического канцерогенеза.
Установлено, что ферменты детоксикации, участвующие в метаболизме проканцерогенов, обнаруживают поразительный полиморфизм. Отдельные изоформы этих белков имеют низкую активность. У индивидуумов с такими вариантами ферментов проканцерогены медленнее подвергаются метаболическим превращениям и выводятся из организма, не успевая превратиться в активные канцерогены. С этим явлением связаны разная чувствительность людей к канцерогенам табачного дыма и предрасположенность курильщиков к раку лёгкого. В покоящихся клетках ДНК двухспиральна, и азотистые основания защищены от воздействия повреждающих агентов. Однако в ходе репликации полинуклеотидные цепи очень чувствительны к канцерогенам, и клетки, получившие повреждения, могут иметь разную судьбу.
Полициклические ароматические углеводороды (ПАУ) входят в состав продуктов неполного сгорания каменного угля и нефти, продуктов пиролиза масел и веществ, найденных в жжёном мясе, а также образуются при курении табака. Они могут связываться с пуриновыми основаниями (особенно гуанином) только после ферментативной активации монооксигеназами, работающими при участии различных изоформ цитохрома Р450. Эти ферменты катализируют образование эпоксидов, которые превращаются в диолы с помощью эпоксидгидролазы. Первичные или вторичные эпоксиды, обладая высокой реакционной способностью, могут взаимодействовать с нуклеофильными группами в молекуле ДНК.
Ароматические амины. К ароматическим аминам относят вещества, использующиеся в производстве анилиновых красителей и резиновой промышленности. Контакт с ними приводит к развитию у рабочих, занятых в указанных производствах, рака мочевого пузыря. Одним из представителей этой группы является 2-нафтиламин, химическая модификация которого происходит главным образом в печени. Канцероген 2-амино-1-нафтол образуется в ходе гидроксилирования 2-нафтиламина. Однако в печени он быстро взаимодействует с ФАФС, превращаясь в нейтральный продукт, который выводится с мочой. В мочевом пузыре часть конъюгатов расщепляется гидролазами, присутствующими в незначительных количествах в моче. Вновь образуется 2-амино-1-нафтол - канцероген, который при повторяющихся контактах человека с нафтиламином вызывает развитие рака мочевого пузыря.
Нитрозамины появляются в организме в результате взаимодействия вторичных алифатических аминов с нитритами. Вторичные амины и нитриты являются постоянными компонентами пищи, поэтому нитрозамины синтезируются при запекании мяса, рыбы. Одно время нитриты широко применялись как консерванты мяса и рыбы, образуются они также в зелёных растениях. Метаболизм нитрозаминов микросомальной системой окисления приводит к образованию иона метилдиазония, который способен метилировать ДНК клеток, индуцируя возникновение злокачественных опухолей лёгких, желудка, пищевода, печени и почек Основным продуктом взаимодействия нитрозаминов с ДНК клетки является N7-метилгуанин-ДНК, но наибольшей канцерогенностью обладает минорный продукт этого взаимодействия - О6-метилированный гуанин-ДНК.
Алкилирующие и ацилируюшие агенты, взаимодействуя с нуклеофильными амино- и гидроксильными группами ДНК, могут повреждать структуру генов и индуцировать образование опухолей. Такие соединения, как винилхлорид, используемый в производстве пластмасс и упаковочных материалов, некоторые лекарства, применяемые в лечении опухолей или как иммуносупрессоры (циклофосфамид, бисульфан, диэтилстильбэстрол), можно рассматривать как факторы риска. Лекарственные препараты этой группы соединений способны вызывать вторичные опухоли у небольшого процента больных.
137. Особенности развития, строения и метаболизма эритроцитов.
Эритроциты - высокоспециализированные клетки, которые переносят кислород от лёгких к тканям и диоксид углерода, образующийся при метаболизме, из тканей к альвеолам лёгких. Транспорт О2 и СО2 в этих клетках осуществляет гемоглобин, составляющий 95% их сухого остатка. Организм взрослого человека содержит около 25×1012 эритроцитов, при этом каждые сутки обновляется примерно 1% этого количества клеток, т.е. в течение одной секунды в кровоток поступает около 2 млн эритроцитов. Эритроциты - единственные клетки, доторые имеют только клеточную мембрану и цитоплазму. Дифференцировка стволовых клеток в специализированные происходит в клетках костного мозга и заканчивается в кровотоке. Особенности строения эритроцитов соответствуют их функциям: большая площадь поверхности обеспечивает эффективность газообмена, эластичная клеточная мембрана облегчает движение по узким капиллярам, специальная ферментативная сисгема защищает эти клетки от активных форм кислорода.
Дифференцировка эритроцитов.Эритроциты, так же как и другие клетки крови, образуются из полипотентных стволовых клеток костного мозга Размножение и превращение начальной клетки эритроидного ряда в унипотентную стимулирует ростовой фактор интерлейкин-3. Интерлейкин-3 синтезируется Т-лимфоцитами, а также клетками костного мозга. Это низкомолекулярный белок группы цитокинов - регуляторов роста и дифференцировки клеток. Дальнейшую пролиферацию и дифференцировку унипотентной клетки эритроидного ряда регулирует синтезирующийся в почках гормон эритропоэтин. Скорость образования эритропоэтина в почках зависит от парциального давления кислорода. При недостатке кислорода скорость образования гормона повышается и, соответственно, количество эритроцитов тоже увеличивается. Хроническая почечная недостаточность сопровождается снижением образования эритропоэтина в почках, что приводит к развитию анемии. В процессе дифференцировки на стадии эритробласта происходят интенсивный синтез гемоглобина, конденсация хроматина, уменьшение размера ядра и его удаление. Образующийся ретикулоцит ещё содержит глобиновую мРНК и активно синтезирует гемоглобин. Циркулирующие в крови ретикулоциты лишаются рибосом, ЭР, митохондрий и в течение двух суток превращаются в эритроциты. Стволовая клетка превращается в эритроцит за две недели. Эритроциты не содержат ядра и поэтому не способны к самовоспроизведению и репарации возникающих в них повреждений. Эти клетки циркулируют в крови около 120 дней и потом разрушаются макрофагами в печени, селезёнке и костном мозге.
Строение эритроцитов.Двояковогнутая форма эритроцитов имеет большую площадь поверхности по сравнению с клетками сферической формы такого же размера. Это облегчает газообмен между клеткой и внеклеточной средой. Кроме того, такая форма, а также особенности строения мембраны и цитоскелета обеспечивают большую пластичность эритроцитов при прохождении ими мелких капилляров. Важную роль в сохранении формы и способности к обратимой деформации эритроцитов играют липиды и белки плазматической мембраны. Липиды бислоя плазматической мембраны эритроцитов, так же, как плазматические мембраны других клеток, содержат глицерофосфолипиды, сфингофосфолипиды, гликолипиды и холестерол. величение содержания холестерола в составе мембраны, которое может наблюдаться при некоторых заболеваниях, снижает её текучесть и эластичность, а следовательно, и способность к обратимой деформации. Это, в свою очередь, затрудняет движение эритроцитов через капилляры и может способствовать развитию гемостаза. Методом электрофореза в мембране эритроцитов обнаруживают около 15 основных мембранных белков с молекулярной массой от 15 до 250 кД. Около 60% массы мембранных белков приходится на спектрин, гликофорин и белок полосы 3 (называется так по расположению этой белковой фракции на электрофореграмме относительно других белков). Интегральный гликопротеин гликофорин присутствует только в плазматической мембране эритроцитов. К N-концевой части белка, расположенной на наружной поверхности мембраны, присоединено около 20 олигосахаридных цепей.
Спектрин - периферический мембранный белок, нековалентно связанный с цитоплазматической поверхностью липидного бислоя мембраны. Он представляет собой длинную, тонкую, гибкую фибриллу и является основным белком цитоскелета эритроцитов. Спектрин состоит из α- и β-полипептидных цепей, имеющих доменное строение; α- и β-цепи димера расположены антипараллельно, перекручены друг с другом и нековалентно взаимодействуют во многих точках. Спектрин может прикрепляться к мембране и с помощью белка анкирина. Этот крупный белок соединяется с β-цепью спектрина и цитоплазматическим доменом интегрального белка мембраны - белка полосы 3. Анкирин не только фиксирует спектрин на мембране, но и уменьшает скорость диффузии белка полосы 3 в липидном слое. Таким образом, на цитоплазматической поверхности эритроцитов образуется гибкая се-тевидная структура, которая обеспечивает сохранение их формы при прохождении через узкие капилляры сосудов Интегральный белок полосы 3 - белок-переносчик ионов С1- и НСО3- через плазматическую мембрану эритроцитов по механизму пассивного антипорта. В разделе 1 подробно описана роль эритроцитов в газообмене. Поступающий из тканей в эритроциты СО2 под действием фермента карбоангидразы превращается в слабую угольную кислоту, которая распадается на Н+ и НСО3-. Образующиеся при этом протоны присоединяются к гемоглобину, уменьшая его сродство к О2, а бикарбонаты с помощью белка полосы 3 обмениваются на Cl- и выходят в плазму крови.
Н2О + СО2 → Н2СО3 → Н+ + НСО3- → обмен на Сl- .
В лёгких увеличение парциального давления кислорода и взаимодействие его с гемоглобином приводят к вытеснению протонов из гемоглобина, обмену внутриклеточного Сl- на НСО3- через белок полосы 3, образованию угольной кислоты и её разрушению на СО2 и Н2О. Мембранный фермент Nа+, К+-АТФ-аза обеспечивает поддержание градиента концентраций Na+ и К+ по обе стороны мембраны. При снижении активности Na+, К+-АТФ-азы концентрация Na+в клетке повышается, так как небольшие ионы могут проходить через мембрану простой диффузией. Это приводит к увеличению осмотического давления, увеличению поступления воды в эритроцит и к его гибели в результате разрушения клеточной мембраны - гемолизу.
Са2+-АТФ-аза- ещё один мембранный фермент, осуществляющий выведение из эритроцитов ионов кальция и поддерживающий градиент концентрации этого иона по обе стороны мембраны.
Метаболизм глюкозы в эритроцитах.Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% - в пентозофосфатном пути. Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа+, К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками - присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином. Глюкоза в эритроцитах используется и в пентозофосфатном пути, окислительный этап которого обеспечивает образование кофермента NADPH, необходимого для восстановления глутатиона.
138. Транспорт кислорода и диоксида углерода кровью. Гемоглобин плода (HbF) и его физиологическое значение.
Кровь осуществляет дыхательную функцию прежде всего благодаря наличию в ней гемоглобина. Физиологическая функциягемоглобина как переносчика кислорода основана на способности обратимо связывать кислород. Поэтому в легочных капиллярах происходит насыщение крови кислородом, а в тканевых капиллярах, где парциальное давление кислорода резко снижено, осуществляется отдача кислорода тканям.
Гемоглобин человека содержит 0,335% железа. Каждый грамм-атом железа (55,84 г) в составе гемоглобина при полном насыщениикислородом связывает 1 грамм-молекулу кислорода (22400 мл). Таким образом, 100 г гемоглобина могут связывать
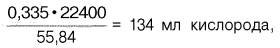
а каждый грамм гемоглобина – 1,34 мл кислорода. Содержание гемоглобина в крови здорового человека составляет 13–16%, т.е. в 100 мл крови 13–16 г гемоглобина. При РО2 в артериальной крови 107–120 гПа гемоглобин насыщен кислородом на 96%. Следовательно, в этих условиях 100 мл крови содержит 19–20 об. % кислорода:

В венозной крови в состоянии покоя РО2 = 53,3 гПа, и в этих условиях гемоглобин насыщен кислородом лишь на 70–72%, т.е. содержание кислорода в 100 мл венозной крови не превышает
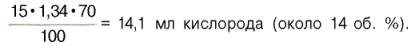
Артериовенозная разница по кислороду будет около 6 об. %. Таким образом, за 1 мин ткани в состоянии покоя получают 200–240 млкислорода (при условии, что минутный объем сердца в покое составляет 4 л). При взаимодействии молекулы кислорода с одним из четырех гемов гемоглобина кислород присоединяется к одной из половинокмолекулы гемоглобина (допустим, к α-цепи этой половинки). Как только такое присоединение произойдет, α-полипептидная цепь претерпевает конформа-ционные изменения, которые передаются на тесно связанную с ней β-цепь; последняя также подвергается конформационным сдвигам. β-Цепь присоединяет кислород, имея уже большее сродство к нему. Таким путем связывание одноймолекулы кислорода благоприятствует связыванию второй молекулы (так называемое кооперативное взаимодействие). После насыщения кислородом одной половины молекулы гемоглобина возникает новое, внутреннее, напряженное состояние молекулыгемоглобина, которое вынуждает и вторую половину гемоглобина изменить конфор-мацию. Теперь еще две молекулы кислорода, по-видимому, по очереди связываются со второй половинкой молекулы гемоглобина, образуя оксигемоглобин.
Организм располагает несколькими механизмами переноса СО2 от тканей к легким. Часть его переносится в физически растворенном виде. Растворимость СО2 в плазме крови в 40 раз превышает растворимость в ней кислорода, тем не менее при небольшой артериовенозной разнице РСО2 (напряжение СО2 в венозной крови, притекающей к легким по легочной артерии, равно 60 гПа, а в артериальной крови – 53,3 гПа) в физически растворенном виде может быть перенесено в покое 12–15 мл СО2, что составляет 6–7% от всего количества переносимого углекислого газа. Некоторое количество СО2 может переноситься в виде карбаминовой формы. Оказалось, что СО2 может присоединяться к гемоглобинупосредством карбаминовой связи, образуя карбгемоглобин, или карбаминогемо-глобин
Карбгемоглобин– соединение очень нестойкое и чрезвычайно быстро диссоциирует в легочных капиллярах с отщеплением СО2. Количество карбаминовой формы невелико: в артериальной крови оно составляет 3 об. %, в венозной – 3,8 об. % . В виде карбаминовой формы из ткани к легким переносится от 3 до 10% всего углекислого газа, поступающего из тканей в кровь. Основная масса СО2 транспортируется с кровью к легким в форме бикарбоната, при этом важнейшую роль играет гемоглобин эритроцитов.
Гемоглобин F — это белок-гетеротетрамер из двух α-цепей и двух γ-цепей глобина, или гемоглобин α2γ2. Этот вариант гемоглобина есть и в крови взрослого человека, но в норме он составляет менее 1 % от общего количества гемоглобина крови взрослого и определяется в 1-7 % от общего числа эритроцитов крови. Однако у плода эта форма гемоглобина является доминирующей, основной. Гемоглобин F обладает повышенным сродством к кислороду и позволяет сравнительно малому объёму крови плода выполнять кислородоснабжающие функции более эффективно. Однако гемоглобин F обладает меньшей стойкостью к разрушению и меньшей стабильностью в физиологически широком интервале pH и температур. В течение последнего триместра беременности и вскоре после рождения ребёнка гемоглобин F постепенно — в течение первых нескольких недель или месяцев жизни, параллельно увеличению объёма крови — замещается «взрослым» гемоглобином А (HbA), менее активным транспортёром кислорода, но более стойким к разрушению и более стабильным при различных значениях pH крови и температуры тела. Такое замещение происходит вследствие постепенного снижения продукции γ-цепей глобина и постепенного увеличения синтеза β-цепей созревающими эритроцитами. Повышенное сродство к кислороду HbF определяется его первичной структурой: в γ-цепях вместо лизина-143 (β-143 лизин у HbA находится серин-143, вносящий дополнительный отрицательный заряд. В связи с этим молекула HbA менее положительно заряжена и основной конкурент за связь гемоглобина с кислородом − 2,3ДФГ (2,3-дифосфоглицерат) — в меньшей степени связывается с гемоглобином, в этих условиях кислород получает приоритет и связывается с гемоглобином в большей степени
139. Полиморфные формы гемоглобинов человека. Гемоглобинопатии. Анемические гипоксии
Дата добавления: 2016-12-26; просмотров: 1022;