Сложные формы обучения общественных насекомых
Естественные или максимальное приближенные к ним модели широко используются при исследовании способности к обучению общественных насекомых, индивидуально-приспособительная деятельность которых поражает своей сложностью и сходством с некоторыми сложными формами обучения позвоночных.
Принципиальное значение для определения роли индивидуумов в жизни семьи общественных насекомых имеет их способность к запоминанию и обучению. Мнения исследователей по этому поводу отражают два принципиально различных подхода к оценке поведения общественных насекомых.
Способность общественных насекомых к обучению и наличие у них некоторых когнитивных процессов признается не всеми исследователями. Д. Мак-Фарленд (1988) скептически относится к существованию таких явлений, сходной точки зрения придерживаются и авторы капитальной сводки "The Ants" (Holldobler, Wilson, 1990). По их мнению, поведение общественных насекомых при обучении очень ригидно, что не позволяет сравнивать их с позвоночными животными. Это заключение базируется на данных, которые свидетельствуют о наличии у общественных насекомых способности к простейшему ассоциативному обучению и до некоторой степени – к латентному обучению, которое происходит, когда фуражир запоминает ориентиры на своем пути. В то же время способность применять полученные навыки в новой ситуации, характерная для позвоночных, у них ограничена. Так, известен опыт, в котором муравьи, научившись преодолевать лабиринт, ведущий к пище, не смогли преодолеть такой же лабиринт, ведущий к гнезду (Weiss, Schneirla, 1967).
Много данных существовало в поддержку другой точки зрения – о наличии у общественных насекомых указанных способностей. У них был, например, продемонстрирован высокий уровень развития памяти. Р. Шовен (1972) писал об экспериментах, показавших, что муравьи помнят пространственно организованную зрительную информацию до 7 месяцев. М. Линдауэр (Lindauer, 1961) выяснил, что пчелы помнят места фуражировки спустя 5 месяцев.
Общественные насекомые, в особенности муравьи, хорошо ориентируются в своей среде обитания. Для такой ориентации, как мы знаем из экспериментов на позвоночных животных, необходима развитая способность не только к запоминанию, но и к формированию пространственного представления, т.е. "когнитивной карты". Эксперименты показали, что память об ориентирах у муравьев обладает определенной гибкостью: разным ориентирам насекомые придают "разное значение", и у них можно даже обнаружить иерархическую организацию системы ориентиров (Длусский, 1967; Кауль, 1983). Например, для муравьев-бегунков (Catagfyphis setypes) более важна форма пищевого ориентира, а затем уже они руководствуются его размерами и цветом. Геометрические признаки ориентира муравьи запоминают быстрее, когда он служит указателем пути к гнезду, а не к пище (Мазохин-Поршняков, Мурзин, 1977). А.В. Сулханов (1979) показал, что у муравьев-рабовладельцев каждый фуражир имеет своеобразную "карту местности", причем наиболее предпочтительной для них является ориентация по точечному источнику света, далее – по цепочке вех и, наконец, по собственному пахучему следу. С муравьями проводили также эксперименты, в которых естественные кормовые участки наполовину или полностью очищали от леса, на территории расставляли искусственные ориентиры, а насекомых подвергали действию искусственного геомагнитного поля (Rosengren, Fortelius, 1986). Оказалось, в противоположность тому, что считалось ранее, что удаленные крупные ориентиры, такие как стволы деревьев, контуры леса и др., в пространственной ориентации муравьев также играют важную роль. Возможно, что многие неудачные лабораторные опыты с использованием мелких ориентиров связаны с игнорированием этой способности.
Способность к ассоциативному обучению общественных насекомых одним из первых исследовал К. фон Фриш (1980). Важным результатом этой работы был вывод, что для суждения о способностях к обучению насекомого важно подобрать методику, адекватную сенсорным и двигательным возможностям вида. Следование этому правилу позволило Т. Шнейрле (Schneirla, 1933) показать, что по скорости обучения ориентации в стационарных лабиринтах муравьи не уступают позвоночным животным. Оказалось также, что спецификой обучения насекомых является его "ступенчатый" характер. Например, в задаче на различение пространственного положения объекта пчелы сначала усваивают различия по признаку "справа-слева", и лишь затем "ближе–дальше" (Мазохин-Поршняков, Семенова, 1987). Особенностью формирования следа пространственной памяти у пчел оказалось и то, что пчелы запоминают окраску нужного им цветка в последние 2 секунды перед тем, как опуститься на него, а расположение наземных ориентиров – только когда улетают от цветка (Menzel, 1968). Таким образом, не только сенсорные возможности, но и особенности формирования памяти в связи со специализацией вида (в данном случае – со спецификой фуражировочного поведения пчелы-сборщицы) сказываются на особенностях процесса обучения.
Очевидно, что особенности мотивационных состояний общественных перепончатокрылых также могут сказываться на специфике обучения и когнитивных процессов. Например, при использовании в процессе обучения таких мотивационных состояний как стремление к возвращению в гнездо (см., например: Карась и др., 1990), наблюдалось более быстрое усвоение навыка. Более того, при разных мотивациях – поиске пищи или поиске входа в гнездо – пчелы и осы приобретают индивидуальный опыт независимо и выбирают в таких ситуациях разные правила поиска (Мазохин-Поршняков, 1968; 1989).
У муравьев обнаружена и способность к переносу навыка. Если обучать муравьев прохождению сложного симметричного лабиринта (Карасьи др., 1990), навык отыскания кратчайшего пути, сформированный в одной половине лабиринта, реализуется при проведении эксперимента в другой его половине.
Таким образом, способность общественных насекомых к ассоциативному обучению не вызывает сомнений, хотя это поведение обладает определенной специфичностью.
В ряде исследований было показано, что эти насекомые обнаруживают и более сложные когнитивные способности (Резникова, 1997). В экспериментах Г.А. Мазохина-Поршнякова (1968, 1989) продемонстрирована способность пчел и ос к обобщению зрительных стимулов, и в конечном итоге – к элементам абстрагирования. Так, насекомое может отличать изображенные на картинке предметы по признаку величины, если в опытах варьировать форму объекта. Можно, например, научить пчелу садиться на самую маленькую фигуру, независимо от ее формы. Более того, пчелы могут научиться различать такие признаки, как "двухцветность" или "парность – непарность расположения фигур". В задачах альтернативного выбора насекомые способны оперировать правилом: "помни, где была  приманка в последний раз, и в следующий раз ищи ее в другом месте" (стратегия win-shift).
приманка в последний раз, и в следующий раз ищи ее в другом месте" (стратегия win-shift).
В опытах Ж. И. Резниковой (1969а, б,; 1983) рыжие лесные муравьи проявили способность усваивать логическую структуру задачи, а также продемонстрировали форму поведения, сходную с экстраполяцией, т.е. предвидением хода событий. Лабораторным семьям муравьев была предложена экспериментальная установка, состоящая из веера полосок ("веток")) прикрепленных к стержню (рис. 4.6.).
Подкреплением служила капля сиропа, которую сначала наносили на конец верхней "ветки". Через каждые 10 минут приманку переносили с ветки на ветку все ниже и ниже, так что путь к ней составлял все больший угол по отношению к исходному направлению. Группа тестированных муравьев уже в ходе первых серий опытов стала "угадывать", где приманка окажется в следующий раз, и ожидать ее на этой ветке. Нужно сказать, что сходные способности демонстрируют и пчелы, которые, следуя указаниям пчелы-разведчицы во время танца (см.: 5.4.3.1), вылетают к указанному ими месту взятка и должны при этом предвидеть путь Солнца по небесному своду.
Следует отметить, что эта задача требует не только способности к экстраполяции, но по своей структуре напоминает другую элементарную логическую задачу, разработанную для изучения рассудочной деятельности позвоночных – тест Ревеша–Крушинского (Крушинский, 1986; Зорина, 1998).
В опытах по изучению сложных форм коммуникации у муравьев (см.: 5.6.4.2) специальная серия была посвящена их интеллектуальным возможностям. Оказалось, что муравьи способны быстро выявлять закономерности в поставленной задаче и использовать их для кодирования, "сжатия" передаваемой информации. Так, если муравей-разведчик передавал фуражирам в гнезде сведения о последовательности поворотов на пути к кормушке в лабиринте "бинарное дерево", то он тратил в 3–5 раз больше времени на передачу информации о случайной последовательности поворотов (например, "направо-налево-налево-направо-налево,."), чем о закономерной последовательности (например, "5раз направо"). Это означает, что и система коммуникации муравьев и их когнитивные способности позволяют им использовать простые закономерности сообщения для более компактной передачи этой информации.
Способность муравьев к подражанию обнаруживается как в виде достаточно простой реакции – следования за разведчиком, обнаружившим пищу, так и в подражании особям другого вида (Reznikova, 1982).
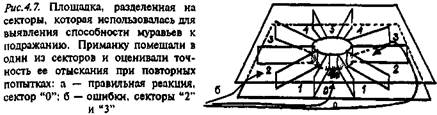
Эксперименты с двумя видами муравьев проводили в естественных условиях, где им предлагали искать пищу в простых экспериментальных установках (рис. 4.7). Фуражиры степного прыткого муравья (Formica cunicularia) выступали в роли "актеров": их обучали находить приманку (см. ниже), фуражиры лугового муравья (F. pratensis) были "зрителями".
В один из таких секторов помещали кусочки приманки. Эти экспериментальные установки размещали на разных площадках. Использовали две схемы опытов: 1) муравьи обоих видов не могли встречаться друг с другом; 2) муравьи-"зрители" имели возможность контакта с "актерами". Попадая в установку впервые, муравьи обнаруживали пищу случайно. При повторных посещениях результаты их поисков могли быть более или менее успешными. Точность усвоения навыка можно было оценить количественно по тому, как далеко ошибочно выбранный муравьем сектор находился от сектора, содержавшего пищу.
Использованные в эксперименте виды муравьев сильно различались и по тактике поведения, и по скорости обучения. Прыткие степные муравьи быстро (через 10–15 мин) находили пищу и уносили ее, "работая" в одиночку. Если муравей этого вида совершал ошибку, т.е. заходил не в тот сектор, то он обегал установку кругом, не заходя в остальные сектора. Луговые муравьи отыскивали пищу очень медленно, а упорядочение поисков при повторных посещениях установки было менее эффективным, чем у другого вида. Совсем иная картина была в случаях, когда луговые муравьи имели возможность наблюдать за поведением особей другого вида. В течение первых 40–50 мин опыта они вообще не трогали пищу, тогда как степные прыткие муравьи (вид "актер") ее растаскивали. Наблюдения показали, что в этих случаях по 3–4 фуражира лугового муравья находились вблизи нужного сектора, наблюдая за действиями фуражиров прыткого степного муравья. Затем они  "актеров" и начинали растаскивать пищу сами. Следует отметить, что при конкуренции за пищу луговые муравьи всегда доминируют над особями второго вида. В таких случаях обучение луговых муравьев отыскиванию сектора с пищей шло гораздо быстрее (рис. 4.8). Эти результаты показывают, что муравьи способны усвоить информацию об оптимальном пути к пище на основе наблюдения за поведением муравьев другого вида.
"актеров" и начинали растаскивать пищу сами. Следует отметить, что при конкуренции за пищу луговые муравьи всегда доминируют над особями второго вида. В таких случаях обучение луговых муравьев отыскиванию сектора с пищей шло гораздо быстрее (рис. 4.8). Эти результаты показывают, что муравьи способны усвоить информацию об оптимальном пути к пище на основе наблюдения за поведением муравьев другого вида.
В заключение следует отметить следующее. Экспериментальные данные, свидетельствующие о способности насекомых к сложным формам обучения и даже к решению аналогов элементарных логических задач, представляются достаточно неожиданными. В настоящее время основные концепции обучения и когнитивной деятельности позвоночных практически не учитывают эту группу фактов. Пока еще не сделано серьезных попыток проанализировать механизмы таких процессов и попытаться объяснить, как они осуществляются в ЦНС насекомых, столь сильно отличной по своему строению от мозга млекопитающих.
4.7.
Этология человека
Широкое применение сравнительного метода к изучению эволюционных основ поведения животных повлекло за собой этологичес-кий анализ видоспецифических элементов в поведении человека. Следует сразу уточнить, что речь идет не о генетических основах интеллекта человека и свойств его высшей нервной деятельности, которым посвящены многочисленные исследования психогенетиков (Равич-Щербо и др., 1999; Малых и др., 1998), а о природе таких реакций как мимические, эмоциональные и некоторые двигательные стереотипы.
Вопрос о существовании наследственно обусловленных элементов в поведении человека долгое время был дискуссионным. По этому вопросу имелся широкий спектр точек зрения. Так, У. Джеймс (см.: 2.3) утверждал, что у человека больше различных инстинктов, чем у животных, но они "замаскированы" его способностью к обучению и мышлению. В отличие от этого большинство бихевиористов придерживалось противоположной точки зрения. Они почти полностью отрицали такую возможность и считали, что ведущую роль в формировании поведения человека играют индивидуально приобретенные компоненты.
Анализ поведения человека с помощью этологических методов одними из первых начали проводить Н. Тинберген (Tinbergen, 1963) и ученик К. Лоренца немецкий ученый И. фон Эйбл-Эйбесфельдт в 50-е годы XX столетия.
Исследование сигнального значения мимических движений человека.Эйбл-Эйбесфельдт исследовал мимику и выражение эмоций, а позднее – особенности восприятия и их проявление в искусстве у представителей ряда этнических групп. В процессе этих исследований он применял те же методы, которые составляют специфику этологии животных. Формулируя цели этого направления науки, он определил, на чем должна базироваться этология человека. Важнейшими направлениями исследования он считал наблюдения за развитием детей в обедненных и обогащенных условиях. Эти данные он планировал сопоставлять с аналогичными, полученными на животных. Важное значение он придавал также сопоставлению наблюдений за особенностями поведения представителей разных культур (кросс-культурный анализ), а также выявлению универсальных филогенетических адаптации в поведении человека, сопоставимых с ФКД, релизерами и другими видоспецифическими феноменами поведения животных.
Благодаря исследованиям, проведенным в разных странах мира (включая Европу, Полинезию и Латинскую Америку), Эйбл-Эйбесфельдт установил, что в поведении человека действительно имеется ряд врожденных двигательных стереотипов, вполне сходных с ФКД животных. К ним относятся в первую очередь многие мимические движения, например вскидывание бровей, знаки согласия и отрицания, поведение при "флирте" и др.
Сравнение поведения представителей разных этнических групп показало, что некоторые из таких стереотипов универсальны и имеют одно и то же значение у народов самых различных культур. Например, "взлет бровей" используется как форма приветствия на расстоянии и у европейских народов, и в Новой Гвинее. Она существует и у японцев, но считается там неприличной и потому активно подавляется воспитанием (Eibl-Eibesfeldt, 1967; 1977). Столь же широко распространенным оказалось и сморщивание носа как гримаса отвращения. Предполагают, что эта реакция возникла в результате эволюционных изменений (ритуализации) непроизвольных движений, совершаемых при рвоте.
 Было обнаружено, что у детей – представителей самых разных этнических групп формирование церемонии приветствия происходит согласно единой стратегии: в них можно обнаружить чередование знаков дружелюбия и агрессивности.
Было обнаружено, что у детей – представителей самых разных этнических групп формирование церемонии приветствия происходит согласно единой стратегии: в них можно обнаружить чередование знаков дружелюбия и агрессивности.
Дата добавления: 2016-08-07; просмотров: 742;