Способность культурных растений выдерживать засоление
| Засоление | Содержание солей, в % от сухой массы почвы | Растения |
| Незначительное | Меньше 0,1 | Все культуры |
| Слабое | 0,1 – 0,4 | Все зерновые, кроме кукурузы; вика, люцерна |
| Среднее | 0,4 – 0,6 | Хлопчатник, тимофеевка, ежа сборная, донник, пшеница |
| Среднесильное | 0,6 – 0,8 | Кормовая брюква, кормовая капуста, пырей, сорго |
| Сильное | 0,8 – 1,0 | Сахарная свекла, подсолнечник, пырей западный, райграс французский, костер безостый |
Повреждающий эффект избыточной концентрации солей объясняется как осмотическими эффектами, так и специфическим влиянием отдельных ионов. Засоление вызывает, прежде всего нарушение водного и минерального обмена растения. Повреждения, вызываемые засолением, сходны с теми, что наблюдаются при засухе. Цитоплазма клеток высших растений обычно содержит 100–200 мМ ионов K+ и 1–10 мМ ионов Nа+. При засолении происходит повышение концентрации натрия и других ионов в цитоплазме, что приводит к инактивации многих ферментов и подавлению синтеза белков.
Растения используют различные способы выживания в условиях засоления. Основной стратегией снижения повреждающего эффекта является изолирование меристем (особенно побеговых) и листьев от воздействия солей в высоких концентрациях. Транспорт солей в растение целиком определяется способностью клеток коры корня противостоять засолению. Плазмалемма клеток корней имеет низкую проницаемость для ионов С1–. Натрий поступает в клетки корней пассивно по градиенту электрохимического потенциала (в отличие от ионов С1–). Процесс активного выведения ионов Nа+ из корня обеспечивают мембранные системы, функционирующие с затратой энергии. Транспорт солей в сосуды ксилемы и далее в надземную часть растения ограничивают пояски Каспари. Поглощаемые соли могут концентрироваться в вакуолях клеток.
У ряда растений для вывода избытка солей имеются специализированные пузыревидные клетки, солевые железки и т. д., с помощью которых растения пытаются избежать накопления солей или приспосабливаются к их высокой концентрации (рис. 7.6).
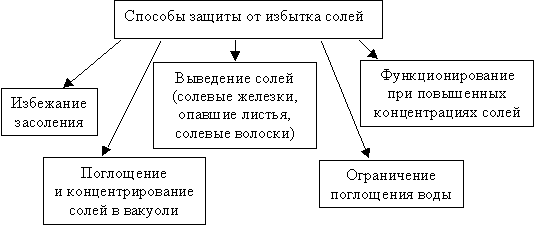
|
| Рис. 7.6. Способы защиты растений от избытка солей |
Растительные организмы способны корректировать водный потенциал клеток при осмотическом стрессе путем дополнительного синтеза таких веществ, как глицинбетаин, пролин, сорбит, сахароза. Количество этих веществ может достигать 10 % веса растения.
При засолении начинают экспрессироваться гены, кодирующие ферменты синтеза пролина и бетаина. Осмотический шок активирует гены, кодирующие ферменты САМ-пути углеродного метаболизма и синтеза лигнина, аквапорины, различные АТРазы и протеазы. В геноме арабидопсиса выявлен локус S0S (salt overly sensitive), который отвечает за ионный гомеостаз и устойчивость к засолению.
Одновременное участие нескольких генов в адаптации к одному стрессовому воздействию свидетельствует о том, что различные гены, отвечающие за устойчивость растений к стрессу, должны иметь общие регуляторные элементы. Изучение промотора стресс-индуцированных генов, экспрессирующихся в ответ на различные стрессовые сигналы, позволило выявить такие нуклеотидные последовательности. Наиболее хорошо изучен ген RD29 (responsive to dehydration), который активируется при осмотическом шоке, охлаждении, водном дефиците и обработке АБК. В промоторной области этого гена выявлены нуклеотидные последовательности, которые позволяют отвечать на АБК (АБК-отвечающий элемент, АВRЕ) и осмотический шок (элемент, отвечающий на дегидратацию, DRЕ).
7.5. Адаптация растений к недостатку кислорода
Ни один из видов растений не способен жить в отсутствии кислорода, но некоторые виды способны переносить более или менее длительный период как недостатка, так и полного отсутствия кислорода.
Корневая система растений обычно получает кислород прямо из почвы. В хорошо дренированных почвах О2 способен диффундировать на глубину до нескольких метров. Поэтому содержание кислорода в газовой фазе почвы практически такое же, как во влажном воздухе. Однако при затоплении и заболачивании корни испытывают недостаток кислорода, тем более, что скорость его диффузии в водной среде невелика. При этом кислород также необходим для дыхания различных других почвенных организмов. Анаэробные условия резко угнетают рост и развитие многих видов растений. Однако некоторые из них (например, рис, ива) очень устойчивы к дефициту кислорода.
Различают условия гипоксии, т. е. частичного дефицита кислорода, и аноксии – полного отсутствия О2 в среде. Большинство растительных клеток и тканей не могут выживать в условиях длительного анаэробиоза. В отсутствие О2 перестает функционировать цикл Кребса и окислительное фосфорилирование. Единственным способом получения АТФ в условиях аноксии является гликолиз. В анаэробных условиях для активного функционирования гликолиза наиболее важен процесс окисления НАДН, который происходит в ходе брожения. При молочнокислом брожении в тканях накапливается молочная кислота (лактат), вызывающая повышение кислотности цитоплазмы (ацидоз). При снижении рН процесс брожения переключается на образование этилового спирта (в кислой среде подавляется активность лактатдегидрогеназы, но возрастает активность лактатдекарбоксилазы). У неповрежденных клеток кислотность вакуолярного содержимого (рН 5,8) гораздо выше, чем цитоплазмы (рН 7,4). При анаэробиозе протоны выходят из вакуоли в цитоплазму и повышают ее кислотность. Именно ацидоз является начальным этапом необратимых нарушений клеточного метаболизма и гибели клетки.
Несмотря на низкую энергетическую эффективность гликолиза, его роль для временного переживания анаэробных условий чрезвычайно велика, особенно, если имеется система детоксикации образующихся при этом продуктов – молочной кислоты и этилового спирта. Детоксикация состоит как в удалении, так и во включении их в обмен веществ. Поэтому способность получать АТФ путем гликолиза и брожения будет зависеть от способности клеток выделять образующийся лактат во внешнюю среду, что позволяет снижать подкисление цитоплазмы. У устойчивых к анаэробиозу растений окисление НАДН, возможно, происходит также за счет обращения дикарбоновой части цикла Кребса в малатдегидрогеназной реакции.
Корни, находящиеся в условиях аноксии или гипоксии, утрачивают способность к эффективному поглощению элементов минерального питания, воды и снабжению ими надземных органов растения. При этом может наблюдаться временное снижение водного потенциала и завядание, которое быстро прекращается, так как вскоре происходит закрывание устьиц, что предотвращает потерю воды за счет транспирации. Однако у некоторых видов растений (горох, томаты) аноксия вызывает закрывание устьиц сразу же, без снижения водного потенциала тканей. Это объясняется тем, что при затоплении так же, как и при дефиците воды, активируется синтез фитогормона АБК и транспорт ее в молодые листья, как из корневой системы, так и из старых подвядших листьев.
Гипоксия индуцирует синтез в корнях предшественника этилена – аминоциклопропанкарбоновой кислоты, которая по ксилеме передвигается в надземную часть, где с участием кислорода превращается в этилен. Именно повышение концентрации этилена является причиной опускания листовых пластинок вниз (эпинастии). Эпинастия наблюдается при попадании корневой системы растения в анаэробные условия.
Водные растения очень хорошо приспособились к постоянному дефициту кислорода в корневой системе. Даже если в воду немного погружены и побеги, они не испытывают видимых признаков стресса. У некоторых видов водных растений, например, таких, как водяная лилия Nутрhoides реltata, при погружении их тканей в воду начинает выделяться эндогенный этилен, который стимулирует интенсивный рост клеток черешка. В результате листья очень быстро оказываются на поверхности воды в воздухе. Сходным путем (с помощью другого фитогормона – гиббереллина) осуществляется регуляция роста междоузлий растений риса при их затоплении. У водного растения рдеста Роtаmоgеtоn ресtinatus, который является обитателем речных, озерных и прудовых водоемов, клетки стебля нечувствительны к этилену, однако их рост при дефиците кислорода также ускоряется за счет подкисления окружающей водной среды СО2, выделяющимся при дыхании.
В стеблях и корнях водных растений, а также растений, приспособленных к условиям затопления, за счет ткани, которая называется аэренхимой, формируется система межклеточных воздушных полостей, пронизывающая практически все растение. По этим воздушным ходам обеспечивается передвижение кислорода и других газов от устьиц до клеток корней. Благодаря такому строению, стебли водных растений значительно легче, чем наземных. Такая же система воздушных полостей образуется (независимо от внешних воздействий), например в растениях риса, что позволяет его корням функционировать в анаэробных условиях на глубине 50 см и более. В корнях риса и других растений, хорошо выдерживающих условия затопления, имеются также структурные барьеры, представленные суберинизированными и лигнифицированными клетками, которые препятствуют диффузии кислорода из тканей корня в почву.
В корнях наземных растений в условиях дефицита кислорода также может формироваться аэренхима (рис. 7.7).

|
| а б |
| Рис. 7.7. Влияние гипоксии на структуру корня кукурузы: а – нормальная аэрация раствора; б – неаэрируемый питательный раствор. Кс – сосуды ксилемы; Эн – эндодерма; Кр – клетки коры; Вп – воздушные полости (аэренхима); Эп – эпидерма |
Этот процесс контролируется фитогормоном этиленом и ионами Са2+. Этилен индуцирует изменения концентрации ионизированного кальция в цитоплазме клеток-мишеней, что запускает механизм программируемой гибели клеток коры корня и образование системы воздушных межклеточных полостей, т. е. аэренхимы. При этом не происходит разрушения клеток центрального цилиндра и эпидермы.
Когда корневая система растений попадает в условия аноксии, синтез большинства белков прекращается, за исключением приблизительно 20 полипептидов. К этим полипептидам, называемым и белками анаэробного стресса, относятся главным образом ферменты гликолиза и брожения. Важным элементом в системе активации генов, продукты которых позволяют растениям адаптироваться к дефициту кислорода, являются колебания концентрации ионизированного кальция в цитоплазме. Одна из наиболее ранних реакций в ответ на аноксию – поступление ионов Са2+ по Са-каналам из митохондрий и некоторых других компартментов в цитоплазму. Уже через несколько минут после воздействия аноксией в цитоплазме клеток кукурузы в культуре регистрируется увеличение содержания ионов Са2+ и количества мРНК, а также, одного из основных ферментов брожения – алкогольдегидрогеназы.
Газоустойчивость
В результате деятельности человека в воздух выделяется более 200 различных газообразных компонентов. Это сернистый газ, оксиды азота, угарный газ, озон, соединения фтора, углеводороды, пары кислот и т. д.
По убыванию токсичности для растений газы располагаются в ряды: F2 > Cl2 > SO2 > NO > CO > CO2; Cl2 > SO2 > NH3 > HCN > H2S.
Пагубное влияние газов на растения проявляется начиная с концентрации 500 мкг/м3.
Газоучтойчивость – это способность растений противостоять действию газов, сохраняя нормальный рост и развитие.
Повреждения различают в зависимости от концентрации газов и по степени усиления повреждений растений отмечают: скрытые, хронические, острые, катастрофические. При действии газов могут наблюдаться такие необратимые явления, как депигментация, некротизация, дефолиация. Следует учитывать также, что при действии нескольких газов создается синергический эффект, т. е. они вызывают более заметные повреждения при более низких концентрациях. Степень отравления газами зависит от различных метеофакторов: температуры, влажности воздуха и почвы, освещенности и т. д.
Косвенное влияние загрязнения приводит к уменьшению плодородия почвы, вызывая гибель полезной микрофлоры, отравление корневой системы, нарушение минерального питания. При понижении рН в результате кислотных дождей увеличивается подвижность токсических металлов. До токсичных концентраций накапливаются, например, цинк, свинец, медь.
Очень сильно страдают от загрязнения хвойные породы: появляется суховершинность, уменьшается длина и увеличивается число хвоинок на побеге, происходит быстрая потеря хвои. У лиственных пород кислые газы вызывают уменьшение размеров и количества листьев, индуцируют появление черт ксероморфности.
Нарушение роста и развития растений под влиянием вредных газов может снижать устойчивость и к другим неблагоприятным факторам среды: засухе, засолению, понижению температуры. Например, растения, выращенные в атмосфере загрязненного воздуха, легче поражаются ржавчиной, серой гнилью, у них снижается интенсивность образования клубеньков.
Различают газочувствительные и газоустойчивые растения. Чуткими индикаторами загазованности служат мхи и лишайники. Отметим, что лишайники устойчивы к обезвоживанию, но не к загрязнению, поскольку у них не сформировалось приспособленности к нему в ходе эволюции.
В результате попадания в атмосферу оксидов серы и азота может происходить закисление осадков и выпадение кислотных дождей. Кислые газы и кислотные дожди нарушают водный режим растений, в результате снижается оводненность тканей, падает содержание связанной воды, изменяется транспирация. Действие газов приводит к закислению цитоплазмы, изменению работы транспортных систем, повреждению плазмалеммы, мембран хлоропластов и других клеточных структур. Вследствие повреждения мембран хлоропластов снижается интенсивность фотосинтеза, разрушается хлорофилл а и каротиноиды, в меньшей степени хлорофилл b и ксантофиллы. Особенно ядовитым для фотосинтеза считается сернистый газ, который лучше растворяется в воде, чем углекислота. Этот газ относят к фотосинтетическим ядам. Интенсивность дыхания вначале повышается, а затем, по мере усиления повреждения и отсутствия поставки субстратов в следствия нарушения фотосинтеза, снижается.
При действии газообразных веществ инактивируется транспорт электронов и всех окислительно-восстановительных ферментов. Нарушается фосфорный обмен, неблагоприятные сдвиги происходят в минеральном обмене. Усиливаются свободно-радикальные процессы, особенно под влиянием озона. В результате фотолиза озона образуются активные супероксид-анион-радикалы, которые могут даже повреждать белки и нуклеиновые кислоты, вызывать мутации, разрушать липиды в результате их перекисного окисления. Озон повреждают мембраны тилакоидов хлоропластов, разрушает РБФ-декарбоксилазу, ингибирует фотосинтез.
Вообще, растения по газоустойчивости делятся на три группы: устойчивые, среднеустойчивые и неустойчивые. Наиболее устойчивые к SO2 древесные породы (вяз, жимолость, лох, клен) оказались устойчивыми и к хлору, фтору, диоксиду азота. Неустойчивыми оказались липа и каштан.
На степень газоустойчивости растений влияет их обеспеченность минеральными элементами. Внесение минеральных удобрений (азот, калий, фосфор) снижает повреждаемость деревьев, особенно липы, каштана, тополя. Замачивание семян в слабых растворах соляной и серной кислот повышает устойчивость растений к кислым газам.
В зависимости от механизмов, ее определяющих, газоустойчивость классифицируется как биологическая, анатомо-морфологическая и физиолого-биохимическая (рис. 7.8).
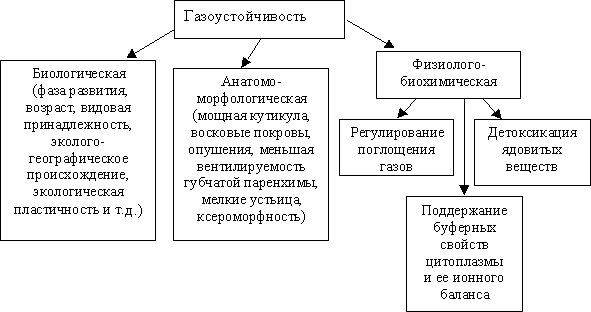
|
| Рис. 7.8. Механизмы газоустойчивости растений |
Биологические механизмы устойчивости связаны с межродовым и межвидовым разнообразием. Например, крестоцветные более устойчивы, чем бобовые, из бобовых фасоль более устойчива, чем клевер, соя и т. д.
Древесные растения (вяз, жимолость, клен) менее устойчивы по отношению к хлору, фтору, закиси азота, чем травянистые. У цветковых повреждаемость листьев зависит даже от их положения на побеге. Эфемеры, отличающиеся интенсивным обменом веществ и коротким вегетативным периодом, относятся к неустойчивым. Виды с более длительным вегетативным периодом лучше переносят условия загазованности. Культурным растениям свойственна большая чувствительность к загрязнению атмосферы по сравнению с дикими видами. Газоустойчивость зависит и от фазы развития, интенсивности роста, возраста растений, положения видов в эволюционной системе, от их эколого-географического происхождения и экологической пластичности.
Анатомо-морфологические признаки, способствующие повышению газоустойчивости, – это мощная кутикула, дополнительные восковые покровы, опушение, меньшая вентилируемость губчатой паренхимы, черты ксероморфности, в частности мелкие устьица. Восковой налет на листьях создает водоотталкивающее покрытие, и грязь легко смывается водой. Восковой налет закрывает также устьичные щели, что повышает устойчивость к загазованности. У С4-растений клетки обкладки проводящих пучков создают барьер, препятствующий воздействию С02 на загрузку флоэмы ассимилятами.
Физиолого-биохимические приспособительные механизмы включают регулирование поступления газов, поддержание буферности цитоплазмы и ее ионного баланса, детоксикацию образующихся ядов. Регуляция поглощения газов определяется, прежде всего, чувствительностью устьиц. Под влиянием газов (особенно сернистого) растения газоустойчивых видов сами закрывают устьица. Например, у растений приспособленных видов при повышении концентрации газов степень открытости устьиц уменьшается на 40 %, а у неприспособленных только на 11 %.
Поддержание ионного баланса и буферных свойств цитоплазмы может быть связано с уровнем в клетках катионов (К+, Nа+, Са2+), способных нейтрализовать ангидриды кислот. Обычно растения, устойчивые к засухе, засолению и некоторым другим подобным воздействиям, имеют более высокую газоустойчивость, возможно благодаря способности регулировать водный режим и ионный состав. На это указывает усиление под влиянием сернистого газа признаков ксероморфности листьев, а под действием хлора признаков суккулентности.
Детоксикация газообразных ядов происходит в результате усиления фитонцидных выделений растений (эфирных масел), обладающих антисептическими свойствами. Это характерно, в частности, для некоторых видов устойчивых к сернистому газу древесных растений. У хвойных повышение уровня сернистого газа приводит к увеличению содержания эфирных масел в хвое. В условиях загазованности подобное обезвреживание ядовитых газов способствует поддержанию фотосинтеза и синтеза в целом на достаточно высоком уровне. У газоустойчивых растений обеспечивается сбалансированность фотосинтеза и оттока ассимилятов. Детоксикация газов может быть результатом и их химического преобразования. Так, в клетках сернистый газ после растворения может дать бисульфит или сульфит. Последний токсичен для растений, но при низкой концентрации метаболизируется хлоропластами до нетоксичного сульфата. При низкой концентрации эффективно обезвреживается и бисульфит, тогда сернистый газ может рассматриваться как источник серы для растения. Однако при повышенных концентрациях этого и других газов их повреждающее действие оказывается настолько сильным, что растения не могут обеспечить их детоксикации.
Уход от воздействия с помощью, например, анатомо-морфологических особенностей – это пассивная устойчивость, а физиологическая способность мириться с поглощением газа или обезвреживать его – активная. Для устойчивых растений характерны: пониженная интенсивность газообмена; высокая скорость метаболизма органических соединений и их транспорта; большая летальная доза при накоплении сернистого газа в клетках и способность к восстановлению обмена веществ, высокая экологическая пластичность. Поэтому в условиях загазованности у приспособленных растений сохраняются необходимый уровень фотосинтеза и дыхания, водный и ионный режимы, катионно-анионный баланс и в целом поддерживается нормальный уровень метаболизма.
Приобретенная газоустойчивость может передаваться по наследству. Например, лиственница японская, растущая вблизи вулканов, более устойчива к сернистому газу, чем европейская, что было установлено после испытания их гибридов. Сопряженная устойчивость проявляется и для газоустойчивых растений: они отличаются большей устойчивостью и к затемнению, засухе, засолению, ионизирующей радиации.
Для повышения газоустойчивости важен отбор видов, устойчивых, в том числе и к другим воздействиям, например к засолению. Повышению уровня устойчивости способствует и выращивание растений на плодородных незагрязненных почвах или смена почвы на газонах и цветниках около промышленных предприятий. Обработка семян слабыми (0,1%-ными) растворами соляной и серной кислот и полив всходов подкисленной водой повышают газоустойчивость, как и периодическое смывание токсических соединений с листьев и нейтрализация известковыми и другими специальными растворами. Эффективны введение физиологически активных соединений, в том числе антиоксидантов – аскорбиновой кислоты, тиомочевины, гидрохинона и др., а также регуляция минерального питания, введение азота в виде мочевины. Несмотря на определенные успехи в исследовании газоустойчивости, общепринятой теории газоустойчивости в настоящее время пока не существует.
Дата добавления: 2016-06-13; просмотров: 1688;