Устойчивость растений к низким температурам
Под устойчивостью к низким температурам подразумевают холодостойкость и морозоустойчивость, т. е. способность растений выживать в условиях низких положительных температур и при заморозках. В листьях растений, повреждающихся при охлаждении, нарушаются процессы фотосинтеза, транспорта ассимилятов, дыхания, синтеза белков. Диапазон повреждающих температур сильно варьирует в зависимости от вида растения. Особенно устойчивы к низким температурам древесные виды в состоянии покоя. Семена, другие обезвоженные ткани растений и споры грибов способны переживать температуры около абсолютного нуля. Тургесцентные вегетирующие клетки также могут выдерживать замораживание, если их охлаждать очень быстро, поскольку формирующиеся в таких условиях кристаллы льда очень малы и не вызывают механических повреждений.
Холодостойкость. Большинство видов тропических и субтропических растений не выдерживают не только заморозков, но даже низких положительных температур. Такие растения, как кукуруза, фасоль, рис, томаты, огурцы и хлопок, останавливаются в росте при температуре 10–15 °С и повреждаются, если температура падает ниже 10 °С. Холодостойкость теплолюбивых растений можно также повысить предпосевным закаливанием. Для этого наклюнувшиеся семена (например, огурцов, томатов) в течение нескольких суток выдерживают в чередующихся (через 12 ч) условиях низких положительных (около 5 °С) и более высоких (10–20 °С) температур. Таким же образом можно закалять и рассаду. Обычное время для закаливания, например, картофеля составляет 15 дней обработки низкими положительными температурами. После такой закалки растения картофеля хорошо переносят даже слабые заморозки. В процессе закаливания изменяется гормональный баланс растения, ростовые процессы затормаживаются, возрастает количество ненасыщенных жирных кислот, включаются механизмы защиты клеток от обезвоживания и замерзания.
Основной причиной повреждающего действия низких положительных температур на теплолюбивые растения является нарушение функционирования клеточных мембран из-за их «затвердевания», связанного с фазовыми переходами жирных кислот, поскольку при достаточно низких температурах липидные бислои ведут себя, как твердые тела. При температуре выше фазового перехода структура бислоя сохраняется, однако при этом жирные кислоты «плавятся», в результате чего вращение и скручивание молекул происходит легче, чем при низких температурах. Затвердевание или «плавление» мембранных липидов зависит от состава содержащихся в них жирных кислот. Для насыщенных жирных кислот с длинной цепью характерны более высокие температуры фазовых переходов. Однако фазовые переходы в мембране индуцируются не только изменениями температуры. Они могут быть вызваны сдвигами рН и мембранного потенциала, двухвалентными катионами и гормонами.
Различная реакция устойчивых и неустойчивых растений на низкие температуры определяется в первую очередь различиями в составе жирных кислот, входящих в состав мембранных фосфолипидов. Выявлено, что у холодостойких растений содержание ненасыщенных жирных кислот (таких, как линоленовая и линолевая) гораздо выше, чем у растений, чувствительных к холоду. Увеличение количества ненасыщенных жирных кислот в составе мембран приводит к снижению температуры фазового перехода мембранных липидов.
У растений, чувствительных к холоду, в составе мембранных липидов велико содержание насыщенных жирных кислот (пальмитиновой, стеариновой). Мембраны такого типа стремятся к затвердеванию до квазикристаллического состояния уже при низких положительных температурах. При этом они становятся менее текучими, что нарушает функционирование многих белков-каналоформеров, переносчиков, рецепторов, ферментов и т. п. Схема влияния низких температур на мембраны от силы воздействия приведена на рис. 7.3.
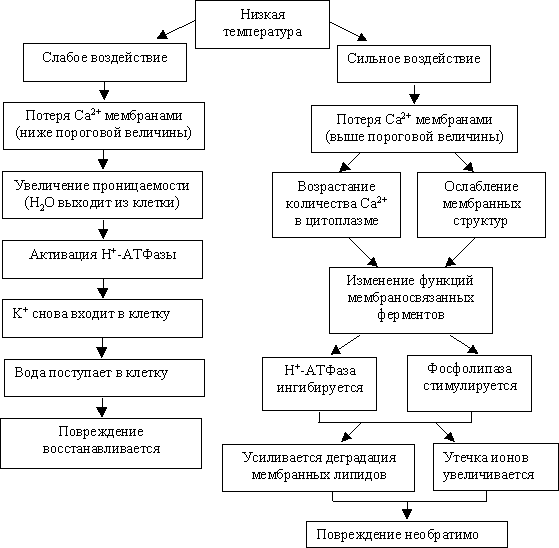
|
| Рис. 7.3. Схема влияния низких температур на мембраны |
Итак, холодостойкими называются растения, которые не повреждаются и не снижают своей продуктивности при температуре от 0 до +10 оС.
Следует отметить, что холодостойкость свойственна и растениям умеренной зоны. Для большинства сельскохозяйственных культур низкие положительные температуры почти безвредны. Наиболее холодоустойчивыми являются яровые пшеница, ячмень, овес. Картофель, томаты, гречиха, просо, кукуруза, соя переносят температуры +5 оС и ниже без значительных повреждений. Фасоль, сорго, клещевина, арахис, рис, арбуз, дыня, тыква, огурцы, кабачки, перец – теплолюбивые растения, погибающие при температуре ниже +10 оС.
В ходе холодового воздействия синтезируются стрессовые белки холодового шока. Так, в проростках озимой пшеницы трехчасовая гипотермия (+3 оС) индуцирует синтез большого набора стрессовых низкомолекулярных и гидрофильных белков.
Морозоустойчивость. Заморозки вызывают повреждение клеток кристаллами льда, образующимися при температуре ниже 0 °С. Повреждающий эффект усугубляется также обезвоживанием тканей, которое при этом происходит. На первых этапах замерзания сильных повреждений растительных клеток не наблюдается, так как вначале кристаллы льда формируются в межклетниках и сосудах ксилемы. Если же заморозки длятся долго, вода переходит из цитоплазмы в апопласт, клетки обезвоживаются, кристаллы льда растут, и повреждают их.
Для предотвращения замерзания в растительных клетках и тканях функционирует система антифризов, представленная различными белками, углеводами и гликопротеинами. Криопротекторы, синтезирующиеся в растительных клетках при понижении температуры, могут предотвратить или резко замедлить рост кристаллов льда. Гидрофильные белки, моно- и олигосахариды, обладающие криопротекторным эффектом, способны связывать значительные количества воды. Связанная таким образом вода уже не замерзает и не транспортируется. Белки и углеводы, обладающие криопротекторным эффектом, способны стабилизировать другие белки и клеточные мембраны при дегидратации клеток, инициированной низкими температурами. Криопротекторы начинают синтезироваться, прежде всего, в эпидермисе и клетках, окружающих межклеточные полости, где происходит наиболее интенсивное образование кристаллов льда при замерзании тканей.
Важная роль в адаптации растений к низким температурам принадлежит фитогормону АБК. Устойчивость многих растений к заморозкам может быть повышена не только закаливанием, но также обработкой их АБК. При воздействии низких положительных температур и (или) при уменьшении светового периода концентрация этого гормона в растительных тканях резко возрастает. Мутанты арабидопсиса, нечувствительные к АБК (abi 1)или с нарушенным синтезом АБК (аbа 1), не могут приспособиться к заморозкам и не закаливаются.
Активность многих генов и белков, которые экспрессируются при низкой температуре или водном дефиците, может быть индуцирована обработкой АБК. При изучении этих генов обнаружилось, что белки, появляющиеся при охлаждении тканей, имеют высокую степень гомологии с семейством белков RAB/LEA/DHN (responsive to ABA, late embryo abudant and dehydrin), синтезируются при созревании семян, водном дефиците и обработке растений АБК. Они очень гидрофильны и обладают необычным свойством: сохраняют свою структуру при резких колебаниях температуры. Продуктами генов LЕА являются гидрофильные белки, способные очень прочно удерживать воду. Считается, что именно эта группа белков обеспечивает защиту клетки при обезвоживании и заморозках, стабилизируя другие белки и клеточные мембраны.
У морозостойких растений при действии низких температур усиливается гидролиз крахмала и накопление в цитоплазме растворимых cахаров. У озимых форм пшеницы и многих других видов растений функцию криопротектора выполняет сахароза, накапливающаяся в больших концентрациях. В качестве антифризов могут также выступать и другие растворимые сахара: рафиноза, фруктозиды, сорбит или маннит, которые накапливаются в клеточных стенках растений и таким образом предотвращают образование кристаллов льда.
Наиболее чувствительны к морозу корни, корневища, клубни и луковицы большинства растений, которые повреждаются при температуре ниже –10 – –15°С. Покоящиеся почки древесных растений способны выдерживать морозы до –25 – –30 °С. У многолетних растений, устойчивых к заморозкам, в период подготовки к зиме в клубнях, луковицах и корневищах накапливаются запасные вещества, которые начинают использоваться с началом вегетационного периода.
Тепловой стресс
При повышении температуры выше оптимальной, при которой физиологические процессы идут с максимальной скоростью, наблюдается их снижение скорости вплоть до остановки.
Только несколько видов высших растений могут постоянно существовать при температуре выше 45 °С. При высоких температурах способны выживать лишь нерастущие и обезвоженные клетки или ткани, например семена и пыльца. Активно вегетирующие ткани редко выживают при температурах более 45 °С, в то время как сухие семена и пыльца некоторых видов могут выдерживать температуру 120 и 70° С соответственно. К высоким температурам хорошо приспособились некоторые суккуленты, для которых характерен САМ-тип углеродного метаболизма. Поскольку у этих растений устьица днем закрыты, они не могут регулировать температуру за счет транспирации, но способны выдерживать перегрев до 60–65 °С. Тепловой стресс представляет потенциальную угрозу при выращивании растений в теплице, где в условиях высокой влажности и неподвижного воздуха плохо функционирует система естественного охлаждения растений путем транспирации.
Одной из основных причин теплового шока, вызываемого высокими температурами, является потеря воды растительными тканями. Вероятно, поэтому продуктивность различных сортов сельскохозяйственных культур (например, хлопка и пшеницы) тем выше, чем больше устьичное сопротивление и соответственно меньше потери воды при транспирации.
При перегреве растительных тканей в первую очередь повреждаются белки и мембраны. Повышается степень ненасыщенности жирных кислот, входящих в состав мембранных липидов, и их текучесть. Снижается способность к образованию водородных и ионных связей между полярными группами биополимеров. Происходит нарушение системы слабых взаимодействий, обеспечивающих поддержание пространственной структурыв белковых молекулах и мембранных липидах. Интегральные мембранные белки стремятся целиком перейти в гидрофобную фазу мембраны, что сказывается на процессах мембранного транспорта, рецепции, энергетике. В результате нарушения третичной структуры элементов электрон-транспортной цепи хлоропластов и митохондрий угнетаются процессы фотосинтеза и дыхания. Наиболее чувствителен к высоким температурам процесс фотосинтеза.
Одним из основных элементов адаптации растительных организмов к перегреву являются белки теплового шока, которые синтезируются при повышении температуры и помогают растениям выдерживать этот тип стрессового воздействия. Впервые они были выявлены у дрозофилы, позже идентифицированы у животных (включая человека), микроорганизмов и растений.
Если клетки или проростки растений быстро нагреть до температуры 40 °С, синтез большинства белков и мРНК будет подавлен. Именно в этих условиях происходит активация синтеза около 30–50 белков, называемых белками теплового шока (БТШ). Новые транскрипты БТШ, т.е. соответствующие мРНК, обнаруживаются уже через 3–5 мин после воздействия высоких температур. Их образование наблюдается также в естественных условиях при постепенном повышении температуры (рис. 7.4).

|
| Рис. 7.4. Влияние низкой температуры на мембраны |
Для растительных организмов характерно накопление низкомолекулярных БТШ (от 15 до 31 кДа). После их синтеза отдельные клетки и целый растительный организм приобретают устойчивость к повышенным температурам и даже к таким, которые ранее были летальными. Нарушения в синтезе БТШ, как правило, лишают растения устойчивости к тепловому шоку. В экспериментах in vitro выявлено, что белковые фракции, обогащенные БТШ с молекулярной массой 15 и 18 кДа, обладают высокой устойчивостью к повышенной температуре и предотвращают денатурацию других белков при нагревании.
Некоторые БТШ не имеют отношения к тепловому шоку, а инициируются другими типами стрессовых воздействий (засухой, низкими температурами, механическими повреждениями, засолением) и обработкой АБК. Таким образом, клетки, подвергшиеся воздействию одного вида стресса, приобретают устойчивость (через образование БТШ) к другим повреждающим факторам, т. е. происходит кросс-адаптация. Например, тепловой шок (48 ч при 38 °С) стимулирует образование ряда БТШ у томатов, что позволяет им без заметных повреждений выдерживать почти 3-недельное пребывание при температуре 2 °С.
Многие БТШ обнаруживаются и в нестрессированных клетках. Различные виды БТШ и их гомологи обнаружены в ядре, митохондриях, хлоропластах, эндоплазматическом ретикулуме, цитоплазме. Выявлено, что БТШ с молекулярной массой 60, 70, 90 кДа и другие работают как молекулярные шапероны, обеспечивая посттрансляционное хранение, созревание (процессинг) и пространственную укладку (фолдиг) единичных белков, а также сборку олигомеров белковых молекул. Некоторые БТШ обеспечивают мембранный транспорт полипептидов в клеточные компартменты и защиту от протеазной активности. Ряд БТШ обладает способностью временно связываться с некоторыми ферментами и высвобождать их только на определенной стадии развития клетки, когда необходимо проявление энзиматической активности.
Дата добавления: 2016-06-13; просмотров: 3058;