Тема 5. Физиология нейронных систем. Рефлексы.
Основное положение рефлекторной теории заключается в утверждении, что деятельность организма есть закономерная рефлекторная реакция на стимул. Узловым моментом развития рефлекторной теории следует считать классический труд И. М. Сеченова (1863) «Рефлексы головного мозга», в котором впервые был провозглашен тезис о том, что все виды сознательной и бессознательной жизни человека представляют собой рефлекторные реакции. Рефлекс как универсальная форма взаимодействия организма и среды есть реакция организма, возникающая на раздражение рецепторов и осуществляемая с участием центральной нервной системы.
В естественных условиях рефлекторная реакция происходит при пороговом, и надпороговом раздражении входа рефлекторной дуги — рецептивного поля данного рефлекса. Рецептивным полем называется определенный участок воспринимающей чувствительной поверхности организма с расположенными здесь рецепторными клетками, раздражение которых запускает рефлекторную реакцию. Рецептивные поля разных рефлексов имеют определенную локализацию, рецепторные клетки — соответствующую специализацию для оптимального восприятия адекватных раздражителей (например, фоторецепторы располагаются в сетчатке; волосковые слуховые рецепторы — в спиральном (кортиевом) органе; проприорецепторы — в мыщцах, в сухожилиях, в суставных полостях; вкусовые рецепторы на поверхности языка; обонятельные — в слизистой оболочке носовых ходов; болевые, температурные, тактильные рецепторы в коже и т. д.
Структурной основой рефлекса является рефлекторная дуга.
Рефлекторная дуга– последовательно соединенная цепочка нервных клеток, которая обеспечивает осуществление реакции в ответ на раздражение.
Рефлекторная дуга состоит из шести компонентов: рецепторов, афферентного (чувствительного) пути, рефлекторного центра, эфферентного (двигательного, секреторного) пути, эффектора (рабочего органа), а также обратной связи.
Рефлекторные дуги могут быть двух видов:
1) простые – моносинаптические (у человека отсутствуют) и дисинаптические рефлекторные дуги. Примером может служить рефлекторная дуга сухожильного рефлекса, состоящая из 3 нейронов (рецепторного (афферентного), вставочного (ассоциативного) и двигательного (эффекторного)), между ними имеется 2 синапса;
2) сложные – полисинаптические рефлекторные дуги. В их состав входят более 3 нейронов – рецепторный, несколько вставочных и эффекторный.
Представление о рефлекторной дуге как о целесообразном ответе организма диктует необходимость дополнить рефлекторную дугу еще одним звеном – петлей обратной связи. Этот компонент устанавливает связь между результатом рефлекторной реакции и нервным центром, который выдает исполнительные команды. При помощи этого компонента происходит трансформация открытой рефлекторной дуги в закрытую и осуществляется контроль точности выполнения реакции.
Рассмотрим свойства полисинаптических рефлексов. В качестве примера полисинаптических рефлексов рассмотрим кашлевой, относящийся к типичным оборонительным. Как известно, ощущение слабого “першения” или “царапания” в горле вызывает кашель, но не сразу, а через короткое время. Задержка связана с тем, что при полисинаптических рефлексах подпороговые стимулы суммируются до надпорогового. Такаясуммация представляет собой центральный процесс: она происходит на уровне интернейронов, а не периферических рецепторов. Неприятные субъективные ощущения (першение, царапание), предшествующие кашлю, убедительно свидетельствуют об опережающем возбуждении рецепторов, ответственных за развитие рефлекса.
По мере увеличения интенсивности стимуляции период времени между началом раздражения (першением) и рефлекторным ответом (кашлем) укорачивается. Иными словами,время рефлексаполисинаптической дугизависит от интенсивности стимуляции: чем она сильнее, тем раньше активируются эффекторы. Уменьшение времени рефлекса объясняется тем, что при росте количества и усилении активности реагирующих на стимул периферических рецепторов быстрее достигается надпороговый уровень возбуждения центральных органов, т. е. время рефлекса уменьшается в первую очередь благодаря временному и пространственному облегчению.
Интенсивность стимуляции влияет и на силу ответа - от легкого покашливания до продолжительного сильного кашля. Это также составляет характерную собенность полисинаптических двигательных рефлексов. Усиление ответа основано на постепенном вовлечении все новых групп мышц - такой процесс названиррадиацией.
Пластичность ответа при полисинаптических рефлексах с пространственно разделенными рецепторами и эффекторами проявляется и в ряде других особенностей, включая “локальный знак”, привыкание (габитуацию), сенситизацию и кондиционирование. Термином“локальный знак” обозначают свойство, иллюстрируемое реакцией на болевое раздражение ноги: степень сокращения сгибателей бедра, колена и стопы зависит от места стимуляции.Привыкание представляет собой ослабление рефлекторного ответа на неболевой и неповреждающий стимул (например, поглаживание кожи живота), который часто повторяется в одном и том же месте с одинаковой интенсивностью. Ослабление ответа не связано с изменениями возбудимости участвующих в рефлексе рецепторов, мотонейронов и скелетных мышц. Изменение места или параметров раздражения (особенно - повышение интенсивности) восстанавливает нормальный ответ, т.е. происходитотвыкание (дизгабитуация). Этому способствует также длительный перерыв в раздражении.
Ритмические болевые стимулы могут приводить ксенситизации. При этом порог рефлекса снижается, его время укорачивается, рецептивное поле расширяется и происходит иррадиация. Терминкондиционирование означает долговременные изменения рефлекторного ответа, обусловленные способностью полисинаптических рефлексов к адаптации и научению. Например, в опыте, когда болевое раздражение можно прекратить только путем движения в сторону стимула, удается вызвать реверсию нормального сгибательного рефлекса.
Рассмотренный кашлевой рефлекс является врожденным. Он наблюдается примерно в одинаковой форме у всех людей. Такие стереотипные реакции в существенной степени предопределены структурной организацией ЦНС. Они возникают автоматически при действии соответствующих стимулов на рецепторы. Например, выделение слюны при попадании пищи в рот, отдергивание руки при уколе пальца и др. Набор врожденных рефлексов одинаков у особей одного вида, поэтому их и называют еще видовыми. Их наличие является таким же обязательным видовым признаком, как форма тела, количество пальцев или рисунок на крыльях бабочки. Для осуществления врожденных рефлексов организм имеет готовые рефлекторные дуги. Центры этих рефлексов расположены в спинном мозге и в стволовой части головного мозга, т.е. в нижних отделах ЦНС. Для их осуществления не обязательно участие коры полушарий большого мозга. Благодаря врожденным рефлексам сохраняется целостность организма, поддерживается постоянство внутренней среды и происходит размножение. Безусловные рефлексы лежат в основе многих поведенческих реакций животных и человека.
Кроме того, каждый организм способен обучаться рефлекторным реакциям, что помогает ему лучше отвечать на постоянно меняющиеся условия внешней среды. Рефлекторные дуги таких приобретенных рефлексов (которые могут снова забываться) находятся на высших уровнях ЦНС. Это, так называемые, приобретенные рефлексы. Они индивидуальны, возникают при определённых условиях и исчезают при их отсутствии, не закрепляются генетически (не передаются по наследству), формируются на базе безусловных рефлексов при участии высших отделов мозга.
Классификации рефлексов. Существуют различные классификации рефлексов: по способам их вызывания, особенностям рецепторов, центральным нервным структурам их обеспечения, биологическому значению, сложности нейронной структуры рефлекторной дуги и т. д.
По способу вызывания различают безусловные рефлексы (категория рефлекторных реакций, передаваемых по наследству) и условные рефлексы (рефлекторные реакции, приобретаемые на протяжении индивидуальной жизни организма).
Различают экстероцептивные рефлексы — рефлекторные реакции, инициируемые раздражением многочисленных экстерорецепторов (болевые, температурные, тактильные и т. д.), интероцептивные рефлексы (рефлекторные реакции, запускаемые раздражением интероцепторов: хемо-, баро-, осморецепторов и т. д.), проприоцептивные рефлексы (рефлекторные реакции, осуществляемые в ответ на раздражение проприорецепторов мышц, сухожилий, суставных поверхностей и т. д.).
В зависимости от уровня активации части мозга дифференцируют спинномозговые, бульварные, мезенцефальные, диэнцефальные, кортикальные рефлекторные реакции.
По биологическому назначению рефлексы делят на пищевые, оборонительные, половые и т. д.
С учетом уровня эволюционного развития, совершенствования сложности нервного субстрата, обеспечивающего соответствующую рефлекторную реакцию, физиологического значения, уровня интегративной деятельности организм а выделяют шесть основных видов рефлексов, или уровней рефлекторных реакций (А. Б. Коган):
Элементарные безусловные рефлексы, представлены простыми рефлекторными реакциями, осуществляемыми на уровне отдельных сегментов спинного мозга. Они имеют местное значение, вызываются локальным раздражением рецепторов данного сегмента тела и проявляются в виде локальных сегментарных сокращений поперечнополосатой мускулатуры. Элементарные безусловные рефлексы осуществляются по жестко детерминированным программам и имеют четкую определенную структурную основу в виде сегментарного аппарата спинного мозга. В результате такие рефлекторные реакции отличаются высокой степенью автоматизма и стереотипности. Функциональная роль этой категории рефлексов заключается в обеспечении простейших приспособительных реакций к внешним воздействиям местного значения, а также в приспособительных изменениях отдельных внутренних органов.
Координационные безусловные рефлексы представляют собой согласованные акты локомоторной деятельности или комплексные реакции вегетативных функциональных объединений внутренних органов. Эти рефлексы также вызываются раздражением определенных групп внешних или внутренних рецепторов, однако их эффект не ограничивается локальной реакцией путем последующей активации широкого класса экстеро-, интеро- и проприорецепторов, а формирует сложные координационные акты сокращения и расслабления, возбуждения или торможения деятельности ряда внутренних органов. В физиологических механизмах реализации рефлекторных реакций этого типа значительное место занимает принцип обратной связи, обеспечиваемый соответствующими спинномозговыми структурами и осуществляющий антагонистическую, реципрокную иннервацию мышц-синергистов и антагонистов. Функциональное назначение координационных безусловных рефлексов — формирование на базе локальных элементарных безусловных рефлексов целостных, целенаправленных локомоторных актов или гомеостатических систем организма.
Интегративные безусловные рефлексы представляют собой дальнейший шаг в интеграции отдельных безусловных рефлексов, осуществляющих сложные двигательные локомоторные акты организма в тесной связи с вегетативным обеспечением, формируя тем самым комплексные поведенческие акты, имеющие определенное биологическое значение. Рефлекторные реакции этого типа инициируются такими биологически важными стимулами, как пищевые, болевые раздражители. Определяющим на входе этих рефлекторных актов становятся не физико-химические свойства стимулов, а в первую очередь их биологическое значение. Интегративные безусловные рефлексы всегда носят целостный системный характер, включая достаточно выраженные соматические и вегетативные компоненты. Их реализация оказывается весьма пластичной, тесно связанной со многими сильно развитыми проприоцептивными обратными связями, обеспечивающими точную коррекцию выполняемого сложного поведенческого акта в соответствии с изменениями в состоянии организма. Пример такой реакции — ориентировочная реакция. Биологическое значение последней заключается в перестройке организма, которая обеспечивает оптимальную подготовку к восприятию и быстрому анализу нового неизвестного сигнала в целях организации рационального ответа. Интегративные безусловные рефлексы требуют для своей реализации надсегментарных механизмов нервно-рефлекторной регуляции поведения организма. Эти рефлексы означают переход от сравнительно простых безусловных рефлексов к поведенческим актам.
Сложнейшие безусловные рефлексы (инстинкты) представляют собой видовые стереотипы поведения, организующиеся на базе интегративных рефлексов по генетически заданной программе. В качестве запускающих стереотипные поведенческие реакции раздражений выступают стимулы, имеющие отношение к питанию, защите, размножению и другим биологически важным потребностям организма.
Сложнейшие безусловные рефлексы образованы последовательными интегративными реакциями, построенными таким образом, что завершение одной реакции становится началом следующей. Адаптивность инстинктов усиливается благодаря наслоению на сложнейшие безусловные рефлексы условных, приобретаемых на ранних этапах онтогенеза. Нервный субстрат, ответственный за физиологические механизмы инстинктивного поведения, представляет иерархическую систему соподчиненных центров интегративных, координационных и элементарных безусловных рефлексов. Жесткая предопределенность инстинктивных реакций обусловлена этапной последовательностью актов инстинктивного поведения, ограничивающей сферу функционирования обратной связи от последующего этапа к предыдущему, уже реализованному. Инстинктивные реакции отражают исторический опыт вида. В субъективной сфере человека сложнейшие безусловные рефлексы проявляются в виде последовательных влечений и желаний, в сложной игре эмоций.
Элементарные условные рефлексы проявляются в интегративных реакциях, вызываемых ранее индифферентными раздражителями, приобретающими сигнальное значение в результате жизненного опыта или подкрепления их безусловными стимулами (сигналами), имеющими биологическое значение. Основным принципиальным отличием этой категории рефлекторных реакций является то, что они образуются в процессе индивидуальной жизни. Условнорефлекторные реакции образуются, усложняются, видоизменяются на протяжении всей жизни; наиболее простые из них формируются в раннем возрасте. Нервным субстратом, отвечающим за осуществление условнорефлекторных реакций, является филогенетически наиболее молодая структура головного мозга — кора больших полушарий. Многоканальность и взаимозаменяемость путей реализации условнорефлекторного механизма лежат в основе высокой пластичности и надежности условнорефлекторных реакций. В системе рефлекторных реакций появление условного рефлекса означает качественно новый скачок в приспособительном поведении высших животных и человека. Условнорефлекторные реакции дают возможность организму заблаговременно отвечать на приближающиеся жизненно важные ситуации. В психической сфере деятельности человека условные рефлексы закладывают начало ассоциативному способу мышления.
Сложные формы высшей нервной деятельности представлены психическими реакциями, возникающими на основе интеграции элементарных условных рефлексов и аналитико-синтетических механизмов абстрагирования. Абстрагирование от конкретного содержания безусловных подкрепляющих раздражителей обеспечивает возможность более полного и целостного восприятия окружающего мира, адекватного прогнозирования и программирования поведения. В качестве вызывающих подобные реакции стимулов обычно выступают сложные комплексные раздражители. Часто такие рефлекторные реакции имеют усеченную рефлекторную дугу (отсутствует эфферентное звено рефлекторной дуги). Сложные формы высшей нервной деятельности оказываются связанными с синтетическими процессами, обеспечивающими целостные субъективные образы внешнего мира, целенаправленные программы поведения, различные проявления абстрактной мыслительной деятельности человека (психонервная деятельность, рассудочное мышление, функции второй сигнальной системы).
Координационная деятельность нервной системы является результатом согласованной работы нейронов, основанной на взаимодействии их между собой. Рассмотрим механизмы работы элементарных нейронных цепей. Они служат для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, обратной связи и др. Они являются основой для выполнения наиболее часто повторяющихся операций.
Особенности взаимодействия афферентных, вставочных и эфферентных нейронов обеспечивают широкий диапазон иррадиации и концентрации возбуждения в ЦНС на основе двух кардинальных принципов: дивергенции и конвергенции.
Дивергенцией называется способность нервной клетки устанавливать многочисленные синаптические связи с различными нервными клетками. Благодаря этому одна нервная клетка может участвовать в нескольких различных реакциях, передавать возбуждение значительному числу других нейронов, которые могут возбудить большее количество нейронов, обеспечивая широкую иррадиацию возбудительного процесса в центральных нервных образованиях.
Рис. Схема дивергенции. Стрелкой показано направление проведения возбуждения по цепочке нейронов.
Процессы конвергенции заключаются в схождении различных импульсных потоков от нескольких нервных клеток к одному и тому же нейрону. Процесс конвергенции характерен не только для однотипных нервных клеток. Частным случаем конвергенции является принцип общего конечного пути. Этот принцип был открыт Ч. Шеррингтоном для мотонейрона спинного мозга. Действительно, активность мотонейрона спинного мозга определяется влиянием со стороны структур самого спинного мозга, различных афферентаций с поверхности тела, посылок от структур ствола, мозжечка, базальных ганглиев, коры мозга и т.д. Полагают, что на поверхности типичного мотонейрона в спинном мозгу человека имеется около 10000 синаптических контактов.

Рис. Конвергенция. Стрелкой показано направление проведения возбуждения по цепочке нейронов.
Реверберация. Итальянский физиолог Лоренто де Но обнаружил наличие в ЦНС замкнутых цепочек нейронов. Попав в такую цепочку, импульс может минутами или часами пробегать небольшой отрезок нейронной цепи, модифицируя при этом синапсы. Сигнал может реверберировать до тех пор, пока или какой-то внешний тормоз не выключит одно из звеньев цепи, или в ней не наступит утомление. Таким образом, при реверберации в цепочке нейронов «хранится» тот сигнал, который включил в ней циркуляцию (реверберацию) импульсов. Считается, что реверберация может служить основой кратковременной памяти.
Большое значение в координации процессов в ЦНС имеет временное и пространственное облегчение (или суммация).
Временное облегчение проявляется в повышении возбудимости нейронов в ходе последовательных ВПСП при действии нескольких следующих друг за другом раздражителей. Механизм заключается в суммации возбуждающих постсинаптических потенциалов.
Пространственное облегчение заключается в возникновение ответной реакции при одновременном действии нескольких подпороговых раздражителей. Например, при раздельной стимуляции двух аксонов возникают подпороговые ВПСП. При совместном раздражении этих аксонов происходит суммация подпороговых ВПСП до надпорогового (возникает ПД). Пространственная суммация имеет большое значение для генерации потенциала действия в клетке.
Пространственная и временная суммации ВПСП способны приводить к длительной деполяризации постсинаптической мембраны и обеспечивать возникновение ритмической импульсной активности нейрона.

Рис. Пространственноеоблегчение и окклюзия
В координационной деятельности центральных нервных образований значительна роль взаимодействия рефлексов, которая проявляется в различных эффектах. Так, феномен суммации можно наблюдать при взаимодействии рефлексов между собой. Противоположное облегчению явление - явление окклюзии.
Окклюзия возникает, например, при одновременном раздражении 2-х афферентных нейронов. При этом ответная реакция может быть меньше арифметической суммы раздражения каждого из них. Механизм: перекрытие синаптических полей, образуемых афферентными частями взаимодействующих рефлексов - в результате импульсы сходятся к одним и тем же нейронам центральной зоны (конвергенция).
В ряде случаев вместо такого ослабления реакции при совместном раздражении рецептивных полей двух рефлексов можно наблюдать феномен облегчения (т. е. суммарная реакция выше суммы реакции при изолированном раздражении этих рецептивных полей). Это результат того, что часть общих для обоих рефлексов нейронов при изолированном раздражении оказывает подпороговый эффект для вызывания рефлекторных реакций. При совместном раздражении они суммируются и достигают пороговой силы, в результате конечная реакция оказывается больше суммы изолированных реакций.
В центральной нервной системе весьма распространен феномен посттетанической потенциации. Он проявляется следующим образом. Раздражая стимулами редкой частоты афферентный нерв, можно получить некоторый рефлекс определенной интенсивности. Если затем этот нерв в течение некоторого времени подвергать высокочастотному ритмическому раздражению (300—400 стимулов в секунду), то повторное редкое ритмическое раздражение приведет к резкому усилению реакции.
Одним из фундаментальных свойств ЦНС является способность к торможению.Торможение– активный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения. Торможение может развиваться только в форме локального ответа. Выделяют несколько видов тормозных сетей или видов торможения:
1. Возвратное торможение
2. Реципрокное торможение
3. Латеральное торможение

Рис. Виды торможения в ЦНС
Возвратное торможение. Для его возникновения необходимо наличие специальных тормозных нейронов – клеток Реншоу. Возвратное торможение – это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам. Возвратное торможение осуществляет торможение нейрона по принципу обратной связи. Нейрон, возбудившись, через коллатерали своего аксона возбуждает тормозные вставочные нейроны. Тормозный нейрон формирует тормозные постсинаптические потенциалы на клетке, которая его активирует. Причем чем более выражен процесс возбуждения в нейроне, тем более выраженным будет и тормозный процесс. Таким образом, с помощью возвратного торможения осуществляется саморегуляция состояния нейрона, автоматически поддерживается определенный уровень его возбудимости. Наиболее ярким примером возвратного торможения является торможение нейронов спинного мозга через тормозные вставочные клетки Реншоу, которые имеют синапсы на этих же нейронах. Что будет, если отключить тормозные влияния на мотонейроны через клетки Реншоу? Известно, например, что столбнячный токсин блокирует передачу тормозного влияния с клетки Реншоу на мотонейрон. Основным признаком столбняка являются судороги, тетанус мышц. Можно сделать вывод, что отключение этой обратной связи приводит к неконтролируемому возбуждению мотонейрона и несанкционированному сокращению (тетанусу) мышц. Таким образом, благодаря активности клеток Реншоу импульсация мотонейронов поддерживается в оптимальном диапазоне, необходимом для управляемого сокращения мышцы. Этот вид торможения есть практически во всех отделах ЦНС.
Реципрокное торможение – это взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов. Этот вид торможения широко представлен в спинном мозге. Примером реципрокного торможения служит торможение мотонейронов, которые управляют мышцами-антагонистами. Известно, что сгибание или разгибание конечностей осуществляется благодаря согласованной работе двух функционально антагонистических мышц: сгибателей и разгибателей. Координация обеспечивается организацией антагонистических отношений между мотонейронами сгибателей и разгибателей, иннервирующих соответствующие мышцы. Реципрокные функциональные отношения складываются в сегментарных структурах спинного мозга благодаря включению в дугу рефлекса дополнительного элемента — специального тормозного нейрона (клетка Реншоу). Схематически в общем виде этот механизм выглядит следующим образом: сигнал от афферентного звена вызывает возбуждение мотонейрона, иннервирующего мышцу-сгибатель (флексор), и через дополнительную клетку Реншоу тормозит мотонейрон, иннервирующий мышцу-разгибатель (экстензор). Так происходит координированное сгибание конечности; напротив, при выполнении разгибания конечности возбуждается мотонейрон мышцы-разгибателя, а через вставочную клетку Реншоу тормозится, угнетается мотонейрон мышцы-сгибателя. Таким образом, акт сгибания конечности, представляющий собой работу мышц-антагонистов, обеспечивается формированием реципрокных антагонистических отношений между нервными центрами соответствующих мышц благодаря наличию специальных тормозных нейронов — клеток Реншоу.
Латеральное торможение. Данный вид торможения широко представлен в структурах нервной системы, имеющих слоистое строение. Латеральное торможение возникает в одном слое клеток. Возбужденный центральный нейрон через коллатерали возбуждает тормозные нейроны, которые образуют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения. Торможение такого рода называют латеральным потому, что образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону. В таких случаях возбуждение направляется только по строго определенному пути. Этот вид торможения распространен на всех уровнях сенсорных систем. Такое торможение наблюдается у соседних элементов сетчатки, а также в зрительных, слуховых и других сенсорных центрах. Латеральное торможение контрастирует (выделяет) существенные сигналы.
Процессы возбуждения и торможения тесно связаны между собой, протекают одновременно и являются различными проявлениями единого процесса. Очаги возбуждения и торможения подвижны, охватывают большие или меньшие области нейронных популяций и могут быть более или менее выраженными. Возбуждение непременно сменяется торможением, и наоборот. Торможение лежит в основе координации движений, обеспечивает защиту центральных нейронов от перевозбуждения. Проявление и осуществление любого рефлекса возможно только при ограничении распространения возбуждения с одних нервных центров на другие.
Нервный центр - сложное объединение нейронов, согласованно включающихся в регуляцию определенной функции или в осуществление рефлекторного акта. Вся деятельность ЦНС является результатом функционирования значительного числа таких центров. Различают чувствительные центры, двигательные центры, центры различных вегетативных функций (дыхания, кровообращения, слюноотделения, глотания, терморегуляции, мочеиспускания и т.д.). В своих высказываниях о пищевом центре И.П.Павлов подчеркивал, что этот центр находится в различных этажах ЦНС и что входящие в него клетки "... есть клетки воспринимающие, так как они воспринимают разнообразные раздражения”. Эти положения Павлова действительны для всех нервных центров. Клетки нервного центра связаны между собой синаптическими контактами и отличаются огромным разнообразием и сложностью внешних и внутренних связей.
В нервных центрах существуют особенности проведения возбуждения:
1. Одностороннее проведение возбуждения - возбуждение передается с афферентного на эфферентный нейрон. Причина: клапанное свойство синапса.
2. Задержка проведения возбуждения: скорость проведения возбуждения в нервном центре на много ниже таковой по остальным компонентам рефлекторной дуги. Чем сложнее нервный центр, тем дольше проходит по нему нервный импульс. Причина: синаптическая задержка. Время проведения возбуждения через нервный центр - центральное время рефлекса.
3. Суммация возбуждения - при действии одиночного подпорогового раздражителя ответной реакции нет. При действии нескольких подпороговых раздражителей ответная реакция есть. Рецептивное поле рефлекса - зона расположения рецепторов, возбуждение которых вызывает определенный рефлекторный акт.
К общим физиологическим свойствам нервной ткани относятся:
1) возбудимость– способность живой ткани отвечать на действие достаточно сильного, быстрого и длительно действующего раздражителя изменением физиологических свойств и возникновением процесса возбуждения.
Мерой возбудимости является порог раздражения. Порог раздражения– это та минимальная сила раздражителя, которая впервые вызывает видимые ответные реакции. Так как порог раздражения характеризует и возбудимость, он может быть назван и порогом возбудимости. Раздражение меньшей интенсивности, не вызывающее ответные реакции, называют подпороговым;
2) проводимость– способность ткани передавать возникшее возбуждение за счет электрического сигнала от места раздражения по длине возбудимой ткани;
3) рефрактерность – временное снижение возбудимости одновременно с возникшим в ткани возбуждением. Рефрактерность бывает абсолютной (нет ответа ни на какой раздражитель) и относительной (возбудимость восстанавливается, и ткань отвечает на подпороговый или сверхпороговый раздражитель);
4) лабильность– способность возбудимой ткани реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом волн возбуждения, возникающих в ткани в единицу времени (1 с) в точном соответствии с ритмом наносимых раздражений без явления трансформации.
Функциональная система– временное функциональное объединение нервных центров различных органов и систем организма для достижения конечного полезного результата. Полезный результат представляет собой жизненно важный адаптивный показатель, который необходим для нормального функционирования организма.
В состав функциональной системы включаются различные органы и системы организма, каждый из которых принимает активное участие в достижении полезного результата.
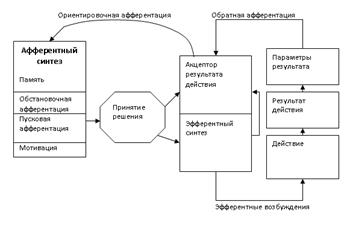
Функциональная система, по П. К. Анохину, включает в себя следующие последовательно сменяющие друг друга стадии:
- афферентный синтез,
- принятие решения,
- акцептор результатов действия,
- эфферентный синтез,
- формирование действия,
- оценка достигнутого результата.
Афферентный синтез включает в себя:
Мотивацию (в ее основе лежит какая-либо потребность); пусковую афферентацию (возбуждения, вызываемые условными и безусловными раздражителями), обстановочную афферентацию (возбуждение от обстановки) и память.
На стадии принятия решения происходит формирование акцептора результата действия (создание идеального образа цели) и формирование программы действия (эфферентный синтез).
Далее следует действие (выполнение программы поведения).
На этапе оценка результата действия идёт сравнение реально выполняемого действия с идеальным образом, созданным на этапе формирования акцептора результата действия (происходит обратная афферентация); на основании результатов сравнения действие или корректируется, или прекращается.
Удовлетворение потребности приводит к прекращению деятельности.
Свойства функциональной системы:
1) Динамичность. В функциональную систему могут включаться дополнительные органы и системы, что зависит от сложности сложившейся ситуации;
2) Способность к саморегуляции. При отклонении регулируемой величины или конечного полезного результата от оптимальной величины происходит ряд реакций самопроизвольного комплекса, что возвращает показатели на оптимальный уровень. Саморегуляция осуществляется при наличии обратной связи.
В организме работает одновременно несколько функциональных систем. Они находятся в непрерывном взаимодействии, которое подчиняется определенным принципам:
1) принципу системогенеза. Происходят избирательное созревание и эволюция функциональных систем (функциональные системы кровообращения, дыхания, питания, созревают и развиваются раньше других);
2) принципу многосвязного взаимодействия. Происходит обобщение деятельности различных функциональных систем, направленное на достижение многокомпонентного результата (параметры гомеостаза);
3) принципу иерархии. Функциональные системы выстраиваются в определенный ряд в соответствии со своей значимостью (функциональная система целостности ткани, функциональная система питания, функциональная система воспроизведения и т.д.);
4) принципу последовательного динамического взаимодействия. Осуществляется четкая последовательность смены деятельности одной функциональной системы другой.
Дата добавления: 2016-05-25; просмотров: 2624;