Тема 4. Физиология межклеточной передачи.
Как передается возбуждение от одного нейрона другому или от нейрона, например, на мышечное волокно? Отросток нейрона - аксон направляется к другому нейрону или исполнительному органу и образует на нем контакт, который называют синапсом (в переводе с греческого - контакт).
Синапс– это структурно-функциональное образование, обеспечивающее переход возбуждения или торможения с окончания нервного волокна на иннервируемую клетку. Один нейрон получает такие контакты, как правило, от нескольких тысяч (3 - 10 тыс.) других нейронов.
Каждый синапс имеет в своем составе:
1) пресинаптическую мембрану (мембрана иннервирующей нервной клетки);
2) постсинаптическую мембрану (мембрана иннервируемой клетки, на которой образован синапс);
3) синаптическую щель (пространство между пресинаптической и постсинаптической мембраной).
Существует несколько классификаций синапсов.
1. По локализации:
1) центральные синапсы (лежат в пределах центральной нервной системы);
2) периферические синапсы.
Центральные синапсы – это контакты между двумя нервными клетками, причем эти контакты неоднородны и в зависимости от того, на какой структуре первый нейрон образует синапс со вторым нейроном, различают:
1) аксосоматический синапс, образованный аксоном одного нейрона и телом другого нейрона;
2) аксодендритный синапс, образованный аксоном одного нейрона и дендритом другого;
3) аксоаксональный синапс - аксон одного нейрона образует синапс на аксоне второго нейрона;
4) дендродентритный синапс - дендрит одного нейрона образует синапс на дендрите другого нейрона.
Различают также несколько видов периферических синапсов:
1) нервно-мышечный синапс, образованный аксоном мотонейрона и мышечной клеткой;
2) нервно-секреторный синапс, образованный аксоном нейрона и секреторной клеткой эндокринной железы.
2. Функциональная классификация синапсов:
1) возбуждающие синапсы;
2) тормозные синапсы.
3. По механизмам передачи возбуждения в синапсах:
1) химические;
2) электрические.
Особенность химических синапсов заключается в том, что передача возбуждения осуществляется при помощи особой группы химических веществ – медиаторов.
Строение химического синапса представлено на рисунке.
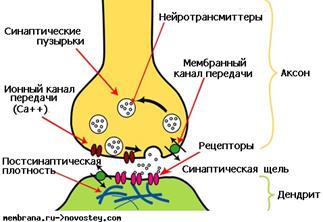
По содержанию медиатора различают несколько видов химических синапсов:
1) холинэргические, в них передача возбуждения происходит при помощи ацетилхолина;
2) адренэргические, передача возбуждения происходит при помощи адреналина или норадреналина;
3) дофаминэргические, передача возбуждения происходит при помощи дофамина;
4) гистаминэргические, передача возбуждения происходит при помощи гистамина;
5) ГАМКэргические, передача возбуждения происходит при помощи гаммааминомасляной кислоты.
Особенность электрических синапсов заключается в том, что передача возбуждения осуществляется при помощи электрического тока. Таких синапсов в организме человека мало.
Разберем сначала каким образом осуществляется передача возбуждения в химическом синапсе? Механизм передачи возбуждения принципиально одинаков во всех химических синапсах. Большая часть того, что известно о синаптических механизмах, получена в экспериментах на определенном синапсе: нервно-мышечном соединении, управляющем сокращением мышц лягушки. Поэтому, для примера, остановимся на механизмах работы именно этого синапса. Процесс начинается с того, что нервный импульс по нервному волокну достигает синапса. Пришедший потенциал действия вызывает деполяризацию мембраны пресинапса. Деполяризация вызывает открытие кальциевых каналов. Ионы кальция из среды, окружающей синапс, входят внутрь пресинапса.
Попадая в цитоплазму синаптического окончания, кальций входит в связь с белками, образующими оболочку синаптических пузырьков, в которых хранится медиатор. Ионы кальция стимулируют движение пузырьков к пресинаптической мембране. Достигая ее, пузырек разрываются, и освобождается ацетилхолин. В каждом синаптическом пузырьке содержится около 10000 молекул ацетилхолина (или 1 квант). Медиатор впрыскивается в синаптическую щель, которая отделяет мембрану одного нейрона от мембраны другого. Этот процесс называется экзоцитозом.
Рис. Схема экзоцитоза.

Таким образом, возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический.
Далее молекулы медиатора начинают диффундировать (двигаться по градиенту концентрации) к постсинаптической мембране. Это довольно медленный процесс, он является причиной синаптической задержки. Достигнув постсинаптической мембраны, ацетилхолин связывается со специальными белковыми молекулами, которые находятся на мембране другого нейрона. Эти молекулы называются рецепторами. Рецепторы устроены уникально и связывают только один тип молекул медиатора, они подходят, как «ключ к замку».
Рецептор состоит из двух частей. Одну можно назвать «узнающим центром», другую - «ионным каналом». Если молекулы медиатора заняли определенные места (узнающий центр) на молекуле рецептора, то ионный канал открывается и ионы начинают входить в клетку (ионы натрия) или выходить (ионы калия) из клетки. Другими словами, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название постсинаптического потенциала.
Постсинаптический потенциал (ПСП) — это временное изменение потенциала постсинаптической мембраны в ответ на сигнал, поступивший с пресинаптического нейрона. Очень важным свойством описанных выше ионных каналов является то, что количество открытых каналов определяется количеством связанных молекул медиатора, а не потенциалом на мембране, как в случае с электровозбудимой мембраной нервного волокна. Таким образом, постсинаптические потенциалы имеют свойство градуальности: амплитуда потенциала определяется количеством молекул медиатора, связанного рецепторами. Благодаря этой зависимости амплитуда потенциала на мембране нейрона развивается пропорционально количеству открытых каналов.

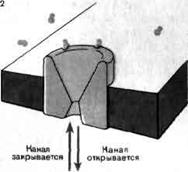

На рисунке схематично изображен ацетилхолиновый рецептор в нервно-мышечном соединении лягушки. Две молекулы ацетилхолина быстро связываются с закрытым в покое каналом и формируют комплекс из рецептора и ацетилхолина (1, 2). Этот комплекс претерпевает конформационные изменения, в результате которых открывается канал для прохождения ионов натрия и калия (3). Время, необходимое для конформационных изменений комплекса, лимитирует скорость реакции. Канал остается открытым в среднем около 1 мс. Пока канал открыт, через него проходит около 20000 ионов натрия и равное количество ионов калия. Ацетилхолин быстро отделяется и разрушается ферментом ацетилхолинэстеразой.
Выделим основные физиологические свойства химических синапсов:
1) способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинаптическую;
2) свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается;
3) свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинаптической задержкой). Это связано с тем, что на пресинаптической и постсинаптической мембране остается медиатор от проведения предыдущего импульса;
4) низкая лабильность синапса (100–150 импульсов в секунду)
5) низкая надежность. Наличие специфических хеморецептивных участков на постсинаптической мембране делает синапсы высокочувствительным к биологически активным веществам. Хеморецептивные зоны часто являются точкой приложения как лекарственных средств, так и различных токсических веществ. Например яд кураре, которым пользовались индейцы Южной Америки при охоте на животных, блокирует рецепторы к ацетилхолину. Это вызывает обездвиживание животного, и оно погибает от удушья.
В состоянии функционального покоя в пресинаптическом окончании происходит случайный контакт синаптических пузырьков с пресинаптической мембраной и выделение в синаптическую щель порции (кванта) медиатора из отдельного пузырька. Выделившийся в синаптическую щель медиатор взаимодействует с хеморецепторами постсинаптической мембраны и приводит к возникновению миниатюрного постсинаптического потенциала.
Синапсы мозга отличаются от синапсов нервно-мышечного соединения в нескольких аспектах. В то время как в нервно-мышечных соединениях ацетилхолин всегда действует возбуждающе, действие того же самого вещества в мозгу в одних синапсах является возбуждающим, а в других тормозным. И если в нервно-мышечных соединениях медиатором обычно служит именно ацетилхолин, то каналы мозговых синапсов регулируются самыми различными медиаторами. Однако каждое данное синаптическое окончание выделяет определенный тип медиатора, и в соответствующей постсинаптической мембране имеются каналы, управляемые этим медиатором. В противоположность активируемым ацетилхолином каналам нервно-мышечного синапса, всегда открывающимся примерно на одну миллисекунду, в некоторых типах мозговых синапсов имеются каналы, открывающиеся на доли миллисекунды, а в некоторых других каналы могут оставаться открытыми сотни миллисекунд.
На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться.
Все эти события происходят на соме и многочисленных отростках дендрита нейрона, на последних находится до нескольких тысяч тормозных и возбудительных синапсов.
В зависимости от характера открытых ионных каналов возникает либо:
1 -возбуждающий постсинаптический потенциал (ВПСП), обеспечивающий деполяризацию постсинаптической мембраны (открываются каналы для ионов натрия и калия), либо
2 - тормозный постсинаптический потенциал (ТПСП), обеспечивающий гиперполяризацию постсинаптической мембраны (открываются каналы для ионов хлора).
ВПСП приближает потенциал клетки к пороговому значению и облегчает возникновение потенциала действия, тогда как ТПСП, напротив, затрудняет возникновение потенциала действия в постсинапсе. Отдельные ПСП обычно невелики по амплитуде и не вызывают потенциалов действия в постсинаптической клетке, однако в отличие от потенциалов действия они градуальны и могут суммироваться.При достижении величины деполяризации мембраны критического уровня генерируется потенциал действия.
Ацетилхолинявляется самым распространенным медиатором в ЦНС и в периферической нервной системе. Также довольно широко распространены в синапсы, в которых медиаторами являются биогенные амины (дофамин, серотонин, адреналин и др.). В этом типе синапсов имеет место следующая последовательность событий. После того как образовался комплекс «молекула медиатора - рецепторный белок», активируется специальный мембранный белок (G-белок). Одна молекула медиатора при связывании с рецептором может активировать много молекул G-белка, и это усиливает эффект медиатора. Каждая активированная молекула G-белка в одних нейронах может открывать ионный канал, а в других активировать внутри клетки синтез специальных молекул, так называемых вторичных посредников. Вторичные посредники могут запускать в клетке многие биохимические реакции, связанные с синтезом, например, белка, в этом случае возникновения электрического потенциала на мембране нейрона не происходит.
В головном мозге в качестве медиаторов «работает» целая группа биогенные амины. В середине прошлого столетия английский врач Паркинсон описал болезнь, которая проявлялась как дрожательный паралич. Это тяжелое страдание вызвано разрушением в мозге больного нейронов, которые в своих синапсах (окончаниях) выделяют дофамин - вещество из группы биогенных аминов. Тела этих нейронов находятся в среднем мозге, образуя там скопление, которое называется черной субстанцией. Исследования последних лет показали, что дофамин в мозге млекопитающих также имеет несколько типов рецепторов (в настоящее время известно шесть типов). Другое вещество из группы биогенных аминов - серотонин (другое название 5-окситриптамин) - вначале было известно как средство, приводящее к подъему кровяного давления (сосудосуживающее). Однако оказалось, что истощение в головном мозге серотонина приводит к хронической бессоннице. В опытах на животных было установлено, что разрушение в мозговом стволе (задних отделах мозга) специальных ядер, которые известны в анатомии как ядра шва, приводит к хронической бессоннице и в дальнейшем гибели этих животных. Биохимическое исследование установило, что нейроны ядер шва содержат серотонин. У пациентов, страдающих хронической бессонницей, также обнаружено снижение концентрации серотонина в мозге.
К биогенным аминам относят также адреналин и норадреналин, которые кроме ЦНС, содержатся также в синапсах нейронов вегетативной нервной системы. Во время стресса под влиянием специального гормона - адренокортикотропного (подробнее см. ниже) - из клеток коры надпочечников в кровь также выбрасываются адреналин и норадреналин.
Медиаторы делят на несколько групп. К классическим нейромедиаторам относят ацетилхолин и биогенные амины, а также аминокислоты: глицин и глутаминовую, аспарагиновую и гамма-аминомасляную (ГАМК) кислоты. Отдельно выделяют нейропептиды: вещество П, энкефалин, соматостатин и др. Медиаторную роль могут выполнять также АТФ, гистамин, пуриновые нуклеотиды.
Согласно функциональной классификации выделяют:
1. Возбуждающие медиаторы, вызывающие деполяризацию постсинаптической мембраны и образование возбуждающего постсинаптического потенциала: ацетилхолин, глютаминовая кислота, адреналин и др.
2. Тормозные медиаторы, вызывающие гиперполяризацию постсинаптической мембраны, после чего возникает тормозной постсинаптический потенциал, который генерирует процесс торможения: ГАМК; глицин и др.
Приняты определенные критерии, по которым вещество относят к группе медиаторов:
1) вещество должно выделяться на пресинаптической мембране в терминали аксона;
2) в структурах синапса должны существовать ферменты, которые способствуют синтезу и распаду медиатора, а также должны быть рецепторы на постсинаптической мембране, которые взаимодействуют с медиатором;
3) вещество, претендующее на роль медиатора, должно при очень низкой своей концентрации передавать возбуждение с пресинаптической мембраны на постсинаптическую мембрану.
В отличие от химического синапса, в электрическом синапсе клетки соединяются высокопроницаемыми контактами с помощью особых белковых туннелей - коннексонов (каждый коннексон состоит из шести белковых субъединиц — коннексинов).

Рис. Строение электрического синапса.
Межмембранные тоннели, сформированные коннексонами, обеспечивают свободный проход через них ионов и малых молекул из одного нейрона в другой. Благодаря прямому переходу ионов через такой синапс нервный сигнал передается ими как по нервному волокну путем возникновения локальных токов.
Рис. Передача возбуждения через электрические синапсы.

Передача возбуждения в электрическом синапсе осуществляется:
- в обе стороны,
- быстрее, чем в электрическом синапсе (без синаптической задержки),
- надежнее.
Электрические синапсы проводят только возбуждающие импульсы, они не утомляемы.
Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические. Электрических синапсов много в раннем онтогенезе (у новорожденных). Во взрослом состоянии они присутствуют в сетчатке глаза и стволе мозга. Обнаружены в небольших количествах также в коре, таламусе, мозжечке и других частях мозга. Функциональное значение электрических синапсов, основанное на быстром проведении ими сигнала в обоих направлениях, заключается в синхронизации активности нейронных популяций. Например, нейронов дыхательного центра ствола головного мозга, генерирующие ритмические электрические импульсы, которые обеспечивают дыхание.
В некоторых синаптических структурах вместе функционируют и электрический и химический механизмы передачи - это так называемые смешанные синапсы.
Дата добавления: 2016-05-25; просмотров: 991;