Тема 3. Физиология нервной клетки
Структурно-функциональной единицей нервной ткани является нервная клетка – нейрон. Нейрон – специализированная клетка, которая способна принимать, кодировать, передавать и хранить информацию, устанавливать контакты с другими нейронами, организовывать ответную реакцию организма на раздражение.
Сколько нервных клеток в нашем мозге? Распространено мнение, что мозг человека состоит из ста миллиардов нейронов: это приблизительно столько же, сколько звезд в нашей Галактике. Число на слуху, правда, никто не знает, откуда оно взялось. В соответствии с имеющимися на сегодня лабораторными данными, в человеческом мозге содержится по разным данным от 15 до 86 миллиардов нейронов.
Формы нейронов чрезвычайно многообразны, не найдется и двух нейронов, одинаковых по виду. Несмотря на это, большинству нейронов присущи определенные структурные особенности, позволяющие выделить три области клетки: клеточное тело, дендриты и аксон. Тело содержит ядро и биохимический аппарат синтеза ферментов и других молекул, необходимых для жизнедеятельности клетки. Обычно тело нейрона имеет приблизительно сферическую или пирамидальную форму. Дендриты представляют собой тонкие трубчатые выросты, которые многократно делятся и образуют ветвистое дерево вокруг тела клетки. Они создают ту основную физическую поверхность, на которую поступают идущие к данному нейрону сигналы. Аксон тянется далеко от тела клетки и служит той линией связи, по которой сигналы, генерируемые в теле данной клетки, могут передаваться на большие расстояния в другие нейроны и исполнительные органы. Аксон отличается от дендритов как по строению, так и по свойствам своей наружной мембраны. Большинство аксонов длиннее и тоньше дендритов и имеет отличный от них характер ветвления: если отростки дендритов в основном группируются вокруг клеточного тела, то отростки аксонов располагаются на конце волокна, в том месте, где аксон взаимодействует с другими нейронами. Нейрон проводит возбуждение только от дендрита к аксону.
Таким образом, основные функции нейрона распределены следующим образом:
1) генерация и передача нервного импульса - передающая часть нейрона (аксональный холмик с аксоном);
2) получение информации - воспринимающая часть (дендриты и мембрана сомы нейрона);
3) способность суммировать возбуждающие и тормозящие сигналы - интегративная часть (тело нейрона).
Основными свойствами нейрона являются раздражимость и возбудимость.
Раздражимость, т.е. способность клетки организма реагировать, то есть изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей. Раздражители – это факторы внешней или внутренней среды, вызывающие раздражение возбудимых структур.
Различают две группы раздражителей:
1) естественные (нервные импульсы, возникающие в нервных клетках и различных рецепторах);
2) искусственные: физические (механические – удар, укол; температурные – тепло, холод; электрический ток – переменный или постоянный), химические (кислоты, основания, эфиры и т.п.), физико-химические (осмотические – кристаллик хлорида натрия).
По биологическому принципу раздражителей делятся на:
1) адекватные, которые при минимальных энергетических затратах вызывают возбуждение ткани в естественных условиях существования организма;
2) неадекватные, которые могут вызвать возбуждение при достаточной силе и продолжительном воздействии.
Второе свойство нейронов, которое обеспечивает нормальное функционирование нервной системы – возбудимость. Возбудимость – свойство клеток отвечать на действие раздражителя специфическими изменениями ионной проницаемости и мембранного потенциала.
Основная роль в процессе возбуждения принадлежит мембране, которая отделяет цитоплазму клетки от окружающей среды. Мембрана нейрона имеет толщину 6-8 нм и состоит из двух слоев липидных молекул, большинство из которых представляет собой сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются повернуты друг к другу – внутрь мембраны, а гидрофильные — наружу в сторону водной среды.
В двойной липидный слой мембраны встроены белки, которые выполняют несколько функций:
1) белки-насосы – пронизывают мембрану насквозь (интегральные белки), перемещают в клетке ионы и молекулы против градиента концентрации;
2) белки-каналы - пронизывают мембрану насквозь (интегральные белки), они образуют места избирательного прохождения для ряда ионов (натрия, калия, кальция, хлора) по градиенту концентрации;
3) рецепторные белки прикреплены на внешней поверхности мембраны и состоят не только из молекул белка, но и из полисахаридов, осуществляют распознавание нужных молекул (например: медиаторов, гормонов и др.) и их фиксацию на мембране;
4) ферменты облегчают протекание химической реакции на внутренней и внешней поверхности нейрона (полуинтегральные белки).
В некоторых случаях один и тот же белок может выполнять функции как рецептора, так и насоса.
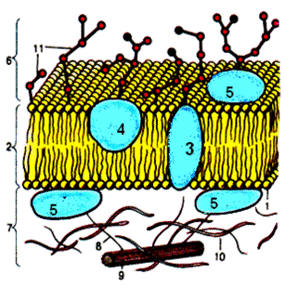
Рис. Схема строения мембраны нейрона:
1. молекула липида;
2. интегральные белки;
3. липидный бислой;
4. полуинтегральные белки;
5. периферические белки;
6. гликокаликс;
7. субмембранный слой;
8. актиновые микрофиламенты;
9. микротрубочки;
10. промежуточные филаменты;
11. углеводные части молекул гликопротеинов и гликолипидов
Основные функции мембраны следующие:
1. Мембрана – граница между внутренней и внешней средой клетки, которые существенно отличаются по своим физико-химическим показателям.
2. Участие в процессах обмена веществ, благодаря наличию ферментов. Большинство биохимических процессов в клетке происходит на мембранах.
3. Регуляция физиологических процессов в клетке. На мембране расположены рецепторные белки, воспринимающие сигналы, поступающие извне и преобразующие их в команды, регулирующие внутриклеточные процессы.
Как же работает нейрон? В любой живой клетке существует разность потенциалов на мембране. Разница разрядов между наружной и внутренней поверхностью покоящейся мембраны называется потенциалом покоя (ПП). ПП нейрона всегда отрицателен, это значит, что внутренняя среда клетки заряжена более отрицательно, чем внеклеточная среда. То есть внутри клетки существует избыточный отрицательный заряд.
Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны. Внутри клетки находится больше всего ионов калия, снаружи его мало. Ионов натрия и ионов хлора больше снаружи, чем внутри. Такое распределение ионов называется ионной асимметрией;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана хорошо проницаема для ионов калия, малопроницаема для ионов натрия и хлора и совсем непроницаема для органических веществ.
За счет чего формируется разность содержания ионов по обе стороны мембраны? Дело в том, что в любой живой клетке в мембране существует постоянно работающий и потребляющий энергию Na+/K+ ионный насос, который откачивает из клетки ионы натрия и закачивает внутрь ионы калия. Существует асимметричность обмена Na+ на K+ в соотношении 3:2. Работа насоса приводит к тому, что внутри клетки концентрация ионов калия примерно в 30 раз превышает их концентрацию вне клетки, тогда как концентрация ионов натрия в клетке примерно в 50 раз меньше, чем снаружи. В результате работыNa+/K+ насоса на мембране появляется слабый электрический потенциала (-10 мВ).
Далее натрий так и остается снаружи (так как мембрана для него малопроницаема), а калий из-за хорошей мембранной проницаемости начинает потихоньку выходить из клетки во внеклеточное пространство. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузии по градиенту. Выходя, калий выносит свой положительный заряд, что приводит к увеличению положительного заряда наружной стороны мембраны (до -70 мВ). Оставшиеся внутри отрицательно заряженные ионы хлора и органические соединения прижимаются к внутренней стороне мембраны (мембрана для них непроницаема). В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны не может быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. В нейроне внутренняя поверхность клетки заряжена отрицательно на величину около 70 мВ относительно внешней.
Измерить этот потенциал можно в лаборатории стеклянным электродом, если в клетку ввести стеклянный микроэлектрод, заполненный раствором соли. Стекло в таком электроде играет роль хорошего изолятора, а раствор соли - проводника. Электрод соединяют с усилителем электрических сигналов и на экране осциллографа регистрируют этот потенциал. Оказывается, потенциал порядка - 70 мВ сохраняется в отсутствие ионов натрия, но зависит от концентрации ионов калия. Другими словами, в создании этого потенциала участвуют только ионы калия, в связи, с чем этот потенциал получил название «калиевый потенциал покоя».
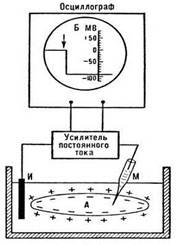
Рис. Регистрация потенциала покоя
Теперь рассмотрим вопрос: Как возбуждается нейрон? Под возбуждением нейрона понимают генерацию нейроном потенциала действия (ПД).
Потенциалом действияназывают сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. Дело в том, что при действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов натрия она повышается в 400–500 раз. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны приобретает отрицательный заряд, внутренняя – положительный.
Как ионные каналы открываются и закрываются? Для каждого типа ионов - натрия и калия - имеется свой собственный тип ионного канала. В покоящемся нейроне натриевые каналы мембраны закрыты и на мембране, как это уже описывалось выше, регистрируется потенциал покоя порядка -70 мВ (отрицательность в цитоплазме). Если же потенциал мембраны деполяризовать (уменьшить поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается. Действительно, в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины. Такой канал называется потенциалзависимым. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией без затраты энергии. Чем больше ионов натрия войдет в цитоплазму нейрона, тем больше его мембрана деполяризуется. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов, возникает лавинообразный ход ионов натрия в клетку. Но потенциал будет расти не бесконечно, а только до тех пор, пока не станет равным примерно +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия, поэтому его называют натриевым равновесным потенциалом. Вспомним, что в покое мембрана имела потенциал -70 мВ, тогда абсолютная амплитуда ПД составит величину около 125 мВ. Мы говорим «примерно» потому, что у клеток разного размера и типов этот потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран.
После того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация»; она наступает примерно через 0,5-1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию-состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (реполяризация). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (-70 мВ) и нейрон готовится к следующему акту возбуждения. Длительность ПД в нервных клетках составляет величину около 1 мс.
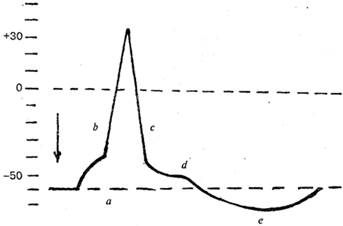
Рис. Потенциал действия: a - локальный ответ; b,c – спайк (b - фаза деполяризации, с - фаза реполяризации); d,e - следовые колебания ( d - отрицательный следовой потенциал; e - положительный следовой потенциал).
Выделяют следующие компоненты потенциала действия:
1) локальный ответ;
2) высоковольтный пиковый потенциал (спайк);
3) следовые колебания:
а) отрицательный следовой потенциал;
б) положительный следовой потенциал.
Локальный ответ связан с небольшим током натрия внутрь клетки. Пока раздражитель не достиг 50–75 % от величины порога, проницаемость клеточной мембраны остается неизменой, и электрический сдвиг мембранного потенциала объясняется раздражающим агентом. Достигнув уровня 50–75 %, открываются потенциалзависимые ворота натриевых каналов, и возникает локальный ответ. Ионы натрия путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это та разность потенциалов, которая должна быть достигнута, чтобы возник лавинообразный ход ионов натрия в клетку.
Высоковольтный пиковый потенциал (спайк).
Пик потенциала действия состоит из двух фаз:
1) восходящей части – фазы деполяризации;
2) нисходящей части – фазы реполяризации.
Лавинообразное поступление ионов натрия в клетку приводит к падению потенциала на клеточной мембране (деполяризации). Чем больше ионов натрия войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется потенциалзависимых натриевых ворот. Постепенно заряд с мембраны снимается, а потом возникает с противоположным знаком. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны или овершутом. Движение ионов натрия внутрь клетки продолжается до момента электрохимического равновесия по иону натрия. Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов натрия и от степени проницаемости мембраны к ионам натрия.
Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. Онахарактеризуется:
1. снижением проницаемости клеточной мембраны для натрия (инактивация натриевых ворот);
2. возрастает проницаемость мембраны для калия, в результате повышается выход калия из клетки с увеличением положительного заряда на мембране;
3. увеличение активности Nа+-К+ насоса, который восстанавливает заряд клеточной мембраны
Однако сразу полного восстановления мембранного потенциала не происходит.
В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный. Следовые потенциалы являются непостоянными компонентами потенциала действия. Отрицательный следовой потенциал результат проникновения внутрь клетки какого-то количества натрия. Таким образом, отрицательный следовой потенциал - это следовая деполяризация.
Положительный следовой потенциал возникает при гиперполяризации клеточной мембраны в процессе восстановления клеточного заряда за счет выхода ионов калия и работы натрий-калиевого насоса. Как только калиевая проницаемость возвращается к исходному уровню - регистрируется мембранный ПП.
Таким образом, нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка - 70 мВ (отрицательность в цитоплазме), а в состоянии возбуждения приобретает потенциал +55 мВ, то есть абсолютная величина потенциала действия - около 125 мВ. Потенциал действия подчиняется закону «всё-или-ничего»: мембрана клетки либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый.
Далее это возбуждение (ПД) должно передаться другому нейрону или какой-то другой клетке, например мышечной, железистой и др. Как же осуществляется проведение возбуждения?
Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки - нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы – миелиновые оболочки, которые образуется из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Через каждые 0,5-1,5 мм оболочка прерывается, формируются перехваты Ранвье. Каждый участок образован только одной клеткой, поэтому перехваты - это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии.
Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения.
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам - кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна.
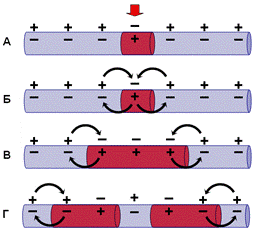
Рис. Схема распространения потенциала действия по немиелинизированному волокну.
Возбужденный участок аксона характеризуется тем, что внутренняя поверхность мембраны, заряжается положительно относительно внешней. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов натрия, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона. На рисунке это отражено стрелками, - выходящий ток деполяризует соседний невозбужденный участок волокна. Возбуждение движется по волокну только в одном направлении и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность - зона невозбудимости. Нам уже известно, что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100 - 120 м/с, должны иметь диаметр около 1 мм.
Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рисунке показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые не покрыта изолятором. Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату – сальтаторно. В узловых перехватах количество натриевых каналов достигает 12 000 на 1 мкм2, что значительно больше, чем в любом другом участке волокна. В результате узловые перехваты являются наиболее возбудимыми. Сальтаторный способ распространения возбуждения экономичен, скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с). Время проведения возбуждения по миелиновому волокну обратно пропорционально длине между перехватами.
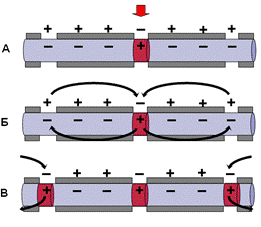
Рис. Распространение потенциала действия по миелинизированным волокнам
Законы проведения раздражения по нервному волокну.
1.Закон анатомо-физиологической целостности.Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
2. Закон изолированного проведения возбуждения. В нервах по нервным волокнам возбуждение передается изолированно.
3. Закон двустороннего проведения возбуждения. Нервное волокно при раздражении проводит нервные импульсы в двух направлениях – центростремительно и центробежно. В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
4. Распространение возбуждения в обе стороны идет с одинаковой скоростью.
5. В миелинизированных волокнах возбуждение проходит, не затухая, без декремента.
Дата добавления: 2016-05-25; просмотров: 889;