Тонус мышц у спинального организма
Спинальный шок. В эксперименте тонус скелетных мышц исследуют после поперечной перерезки спинного мозга (рис. 49).
Сразу после перерезки или повреждения спинного мозга у человека в случае травмы наблюдаются мышечная атония и отсутствие рефлексов (спинальный шок). Главной причиной спинального шока является выключение влияния вышележащих отделов ЦНС на спинной мозг. Об этом свидетельствует тот факт, что повторная перерезка спинного мозга ниже первой повторно шок не вызывает. Спинальный шок у лягушек длится несколько минут, у собак — несколько дней, у человека — около 2 мес. После исчезновения спинального шока тонус мышц, иннервируемых посредством сегментов спинного мозга, которые находятся ниже перерезки (повреждения), резко повышается.
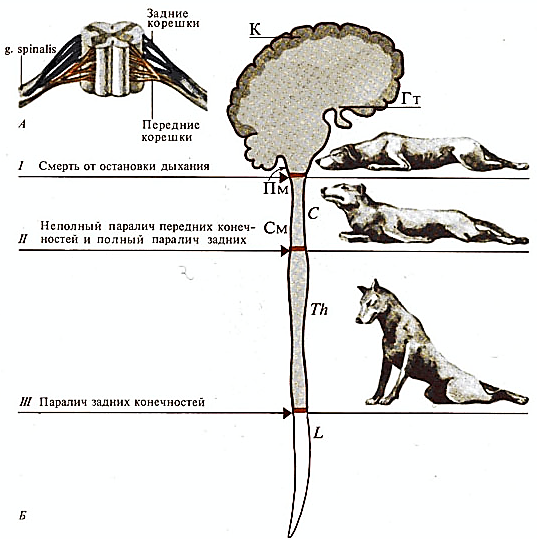
Рис. 49. Последствия перерезки спинного и продолговатого мозга у собаки. Перерезка спинного мозга на различных уровнях вызывает нарушение тонуса и паралич конечностей, а перерезка на уровне продолговатого мозга вызывает остановку дыхания и смерть животного. В продолговатом мозге расположен жизненно важный дыхательный центр.
Гипертонус мышц спинального организма. Сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются (рис. 50).

Рис. 50. Механизм регуляции тонуса мышц на уровне спинного мозга и развития гипертонуса мышц нижних конечностей в случае повреждения спинного мозга: 1 – полусегмент спинного мозга; 2 – скелетная мышца; 3 – мышечное веретено; α – мотонейрон, иннервирующий миоциты скелетной мышцы; γ – мотонейрон, иннервирующий миоциты интрафузального мышечного волокна (проприорецептора)
Гипертонус имеет рефлекторную природу: деафферентация, например, нижней конечности у лягушки ведет к исчезновению тонуса у нее (опыт Бронжеста). Однако произвольные движения невозможны! Афферентная импульсация от мышечных рецепторов по задним корешкам спинного мозга (рис. 50), которая ведет к возбуждению α-мотонейронов спинного мозга и сокращению скелетных мышц и развитию гипертонуса, поддерживается двумя способами: 1) спонтанной активностью мышечных рецепторов, что обеспечивает поступление импульсов от них к α-мотонейронам, возбуждение последних и сокращение мышц; 2) спонтанной активностью γ-мотонейронов.
Возбуждение γ-мотонейронов вызывает возбуждение и сокращение иннервируемых ими интрафузальных мышечных волокон, в результате чего увеличивается натяжение мышечного рецептора, поскольку концы его зафиксированы на скелетной мышце. Это вызывает раздражение и возбуждение мышечных рецепторов, импульсы от которых поступают к α-мотонейронам и возбуждают их.
В свою очередь, α-мотонейроны посылают импульсы к скелетной мышце и вызывают ее постоянное (тоническое) сокращение, вследствие чего и развивается гипертонус у спинального организма. Следует отметить, что в нормальных условиях деятельность спинного мозга контролируется вышележащими отделами ЦНС, нисходящие влияния которых являются преимущественно тормозными.
Ствол мозга
Ствол мозга включает продолговатый мозг, мост и средний мозг, где локализуются ядра черепных нервов продолговатого мозга (IX-XII нервов), моста (V— VIII пары) и среднего мозга (III, ГУ пары), а также другие ядра, управляющие мускулатурой туловища и конечностей.
Функции отдельных ядер черепных нервов рассмотрим в восходящем ряду: продолговатый мозг, мост и средний мозг (рис. 51).
Двигательное ядро добавочного нерва (XI, п. accessorius, продолговатый и спинной мозг), иннервируя грудиноключичную и трапециевидную мышцы, обеспечивает их сокращение и поднимание плечевого пояса кверху, лопаток — к позвоночнику, повороты и наклоны головы.
Ядра языкоглоточного нерва (IX, п. glossopharyngeus, продолговатый мозг). Чувствительное ядро одиночного пути формирует все виды чувствительности слизистой оболочки глотки, задней трети языка, барабанной полости и каротидного тельца, участвует в рефлексах жевания, глотания, в секреторных и моторных пищеварительных рефлексах, в сосудистых и сердечных рефлексах (из каротидного тельца). Нижнее слюноотделительное (парасимпатическое) ядро стимулирует секрецию околоушной слюнной железы. Двойное (двигательное) ядро обеспечивает поднимание глотки и гортани, опускание мягкого нёба и надгортанника в глотательном рефлексе.
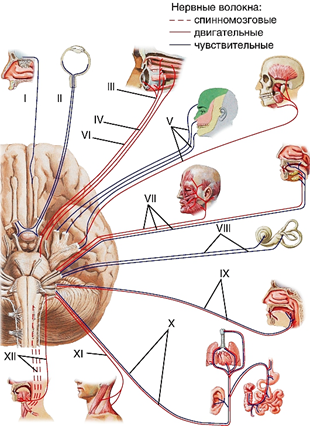
Рис. 51. Черепные нервы — 12 парных нервов, отходящих от головного мозга. I — обонятельный нерв; II — зрительный нерв; III — глазодвигательный нерв; IV — блоковый нерв; V — тройничный нерв; VI — отводящий нерв; VII — лицевой нерв; VIII — преддверно-улитковый нерв; IX — языкоглоточный нерв; X — блуждающий нерв; XI — добавочный нерв; XII — подязычный нерв. Три черепных нерва являются чувствительными (I, II, VIII); шесть — двигательными (III, IV, VI, VII, XI, XII) и три — смешанными (V, IX, X) (Неттер, 2003)
Двигательное ядро подъязычного нерва (XII, п. hypoglossus, продолговатый мозг) иннервирует мышцы языка и обеспечивает его движение в процессе разговора, жевания, глотания, сосания.
Ядра преддверно-улиткового нерва (VIII, п. vestibulocochlearis, продолговатый мозг) — чувствительные. Улитковые ядра несут информацию от слуховых рецепторов, входят в состав слуховой сенсорной системы. Вестибулярные ядра воспринимают информацию от рецепторов вестибулярного аппарата.
Ядра блуждающего нерва (X, п. vagus, продолговатый мозг). Заднее (парасимпатическое) ядро регулирует функции внутренних органов. Чувствительное ядро одиночного пути, иннервирующее слизистую оболочку нёба, корня языка, дыхательных путей, аортальное тельце, органы шеи, грудной и брюшной полости, участвует в качестве афферентного звена в глотательном, жевательном, дыхательных, висцеральных рефлексах. Двойное (двигательное) ядро обеспечивает сократительную активность мышц нёба, глотки, верхней части пищевода и гортани, реализует рефлексы глотания, рвоты, формирование голоса, кашля, чиханья.
Ядра лицевого нерва (VII, п. facialis, мост). Двигательное ядро вызывает сокращения мимической и вспомогательной жевательной мускулатуры, регулирует передачу звуковых колебаний в среднем ухе в результате сокращения стремянной мышцы. Чувствительное ядро одиночного пути, иннервируя вкусовые луковицы передней у языка, участвует в анализе вкусовых раздражителей, в осуществлении моторных и секреторных пищеварительных рефлексов. Верхнее слюноотделительное (парасимпатическое) ядро стимулирует выделение секретов подъязычной, подчелюстной слюнных желез и слезной железы.
Ядро отводящего нерва (VI, п. abducens, мост) обеспечивает сокращение наружной прямой мышцы глаза, в результате чего он поворачивается наружу.
Ядра тройничного нерва (V, п. trigeminus) включают три чувствительных (среднемозговое, мостовое, спинномозговое) ядра и одно двигательное. Среднемозговое чувствительное ядро получает импульсацию от мышц языка, жевательных и мышц глазных яблок. Мостовое и спинальное чувствительные ядра получают тактильную, температурную, висцеральную и болевую импульсацию от первичных афферентных нейронов, расположенных в тройничном ганглии и иннервирующих кожу, слизистые оболочки, органы лица и головы. Информация, поступающая от них, обеспечивает формирование соответствующих ощущений и поведенческих реакций. Двигательное ядро тройничного нерва управляет главными жевательными мышцами; мышцами, напрягающими мягкое нёбо и барабанную перепонку; афферентную импульсацию оно получает от сенсорных ядер V нерва и нейронов РФ.
Ядра глазодвигательного нерва (III, п. oculomotorius, средний мозг). Двигательное ядро обеспечивает сокращение верхней, нижней и внутренней прямой, нижней косой мышц глаза и мышцы, поднимающей верхнее веко. Добавочное (парасимпатическое) ядро, иннервируя ресничную мышцу и сфинктер зрачка, осуществляет соответственно рефлексы аккомодации глаза и сужения зрачка.
Ядро блокового нерва (IV, n. trochlearis, средний мозг), иннервируя верхнюю косую мышцу, осуществляет поворот глаза кнаружи и вниз.
Центры ствола мозга целесообразно объединяют в три группы.
1. Жизненно важные вегетативные центры регуляции систем: дыхания, сердечно-сосудистой системы, пищеварения. Поражение продолговатого мозга ведет к остановке дыхания и смерти.
2. Центры защитных рефлексов, в осуществлении которых участвуют по несколько черепных нервов, — это рефлексы чиханья, кашля, мигания, слезоотделения, рвоты.
3. Ядра, управляющие мускулатурой конечностей и туловища (рис. 52).
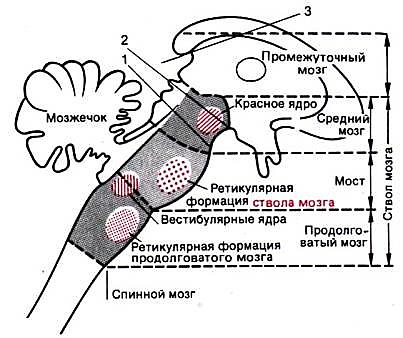
Рис. 52. Схема расположения двигательных центров в стволе мозга (продолговатом мозгу, мосте и среднем мозгу). Нарушая связь с отделами мозга, расположенными выше каждой из трех указанных линий перерезки, получают соответственно децеребрированное (1), мезенцефальное (2) и таламическое (3) животное (Шмидт, 2005)
Ядро шва (продолговатый мозг). Аксоны его нейронов поступают в спинной мозг, многие из них содержат серотонин. На интернейронах заднего рога этот путь образует тормозные синапсы (угнетение передачи сигналов), а на мотонейронах — возбуждающие. В целом, моноаминонергические пути модулируют реактивность нейронных цепей спинного мозга, а не конкретные движения.
Вестибулярные ядра (мост) оказывают возбуждающее влияние на α- и γ-мотонейроны спинного мозга мышц-разгибателей конечностей, туловища, шеи и тормозное влияние на мотонейроны сгибателей этих частей тела. Подобный эффект вызывают и ретикулярные ядра моста (медиальный ретикулоспинальный путь).
Красные ядра (средний мозг, руброспинальный путь), напротив, оказывают тормозное влияние на мышцы-разгибатели и возбуждающее — на мышцы-сгибатели. Такое же влияние оказывает и ретикулярное гигантоклеточное ядро продолговатого мозга (рис. 53).

Рис. 53. Структурные компонеты среднего мозга (поперечный срез). СВ – сильвиев водопровод, БЧ – бугры четверохолмия, РФ – ретикулярная формация, ЧС – черная субстанция, КЯ – красное ядро
Статические и статокинетические рефлексы, замыкающиеся на уровне ствола мозга, регулируя тонус мышц, обеспечивают естественную позу, восстановление нарушенной позы и сохранение равновесия при перемещении организма в пространстве (Магнус Р., 1924).
Статические рефлексы обеспечивают естественную позу в покое и восстановление нарушенной позы. Их разделяют на рефлексы позы (положения тела, с вестибулорецепторов) и выпрямительные рефлексы. Шейные рефлексы позы рассматривались выше.
Вестибулярные рефлексы позы направлены на сохранение естественной позы, они замыкаются на уровне моста. Для их выявления на шею животного накладывают гипсовую повязку, чтобы исключить рефлексы с проприорецепторов шеи. При естественном положении головы импульсация с рецепторов лабиринта минимальная. Если животное располагается спиной вниз, то тонус мышц-разгибателей становится максимальным (лабиринтный рефлекс); если животное располагается спиной вверх — минимальным. При вертикальной позе человека (голова вверх, шея несколько наклонена вперед) шейные и лабиринтные рефлексы усиливают друг друга, происходит повышение тонуса разгибателей нижних конечностей и сгибателей верхних конечностей. После выключения вестибулярного аппарата в эксперименте у животного вестибулярный рефлекс исчезает (опыт Р. Магнуса). Этот рефлекс сохраняется у бульбарного организма (перерезка ствола мозга между мостом и верхними бугорками четверохолмия среднего мозга), однако возникает резкое преобладание тонуса мышц-разгибателей и понижение тонуса мышц-сгибателей — децеребрационная ригидность (Шеррингтон Ч., 1896) (рис. 54).
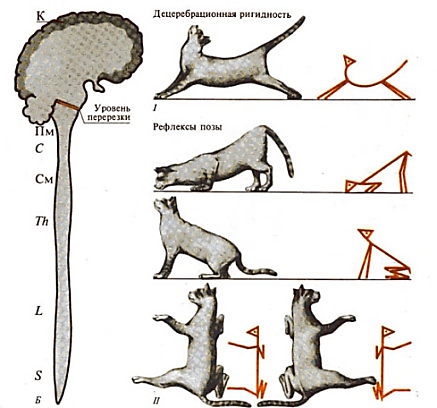
Рис. 54. Развитие децеребрационной ригидности при перерезке головного мозга между мостом и средним мозгом (Чеснокова, 2007)
Ригидность развивается вследствие преобладания возбуждающих нисходящих влияний на α- и γ-мотонейроны над тормозными (см. рис. 55).
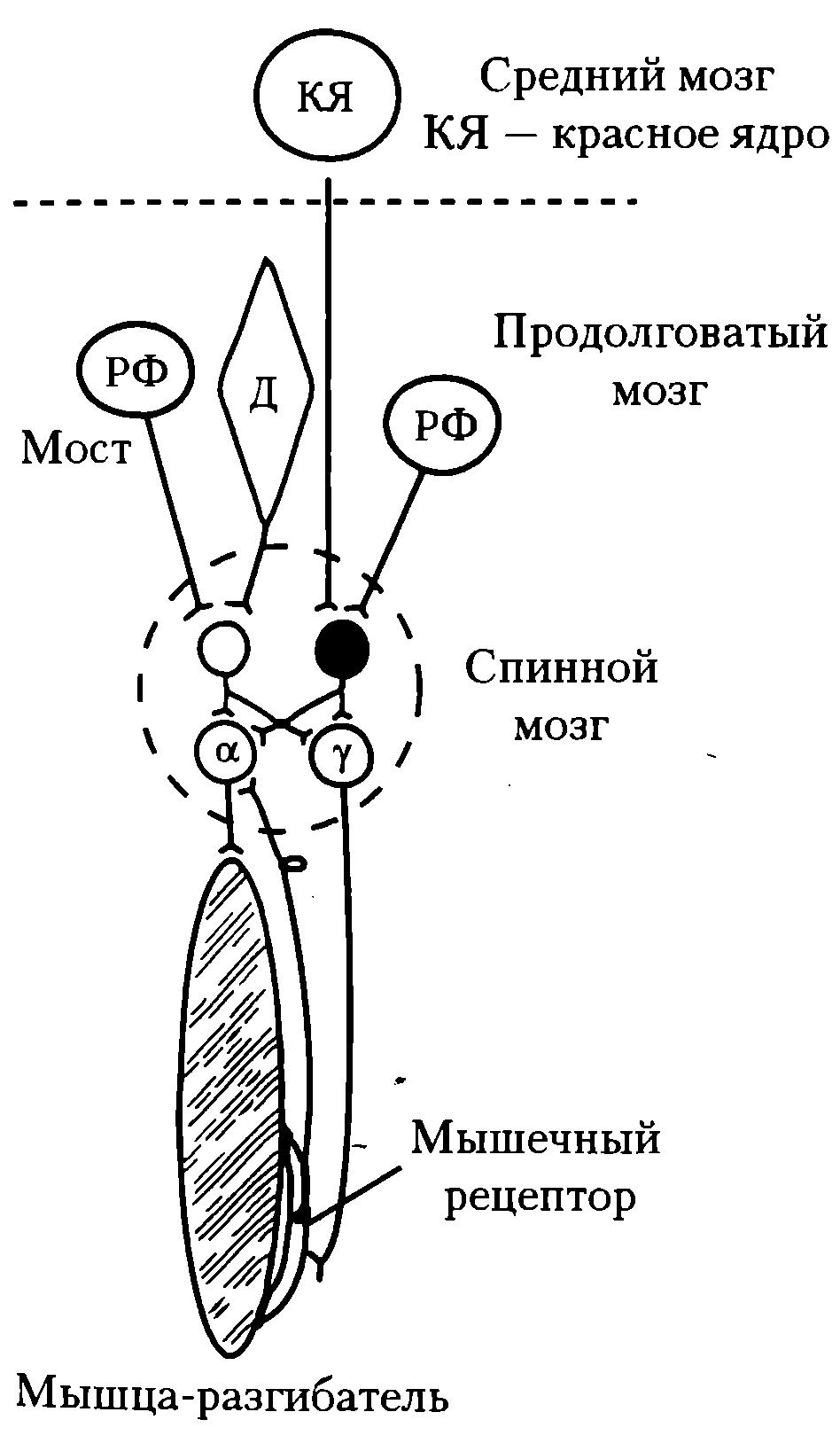
Рис. 55. Механизм регуляции тонуса мышц и возникновения децеребрационной ригидности при перерезке ствола мозга между средним мозгом и мостом (пунктирная линия): РФ – ретикулярная формация; Д – ядро Дейтерса; α- и γ – мотонейроны; вставочные нейроны спинного мозга: светлый – возбуждающий, черный - тормозной
Бульбарный организм не может поддерживать естественную позу. При наклоне головы влево или вправо повышается тонус мышц-разгибателей на соответствующей стороне (предотвращение падения вследствие смещения центра тяжести). При поражении лабиринта одного уха человек часто падает на эту сторону.
Выпрямительные рефлексы наблюдаются у мезэнцефального организма при обязательном участии красного ядра (перерезка в эксперименте между средним и промежуточным мозгом). Благодаря этим рефлексам организм способен принимать естественную позу при ее нарушении, например, при положении животного на боку. Вначале выпрямляется голова (рефлексогенная зона — вестибулярный аппарат и кожа), затем — туловище (рефлексогенная зона — проприорецепторы мышц шеи и рецепторы кожи). При выключении этих рецепторов рефлексы исчезают. В условиях натуральной жизни важную роль в осуществлении выпрямительных рефлексов играет зрение.
Статокинетические рефлексы возникают при ускорениях прямолинейного или вращательного движений организма. Сокращения мышц при этом направлены на преодоление действующих на человека ускорений, на сохранение нормальной позы, ориентации в пространстве. Они также осуществляются с помощью среднего мозга. Эти рефлексы запускаются с рецепторов вестибулярного аппарата. Они включают лифтные рефлексы, нистагм головы и глаз, перераспределение мышечного тонуса при ходьбе и беге.
Лифтные рефлексы. При остановке движущегося вниз лифта и в начале быстрого подъема лифта, в нижних конечностях повышается тонус мышц-разгибателей, что обеспечивает преодоление сил инерции, хотя наблюдается некоторое сгибание конечностей под влиянием массы тела. В начале опускания лифта и при остановке движущегося вверх лифта наблюдаются противоположные явления. Правда, существует и другая точка зрения, учитывающая только внешние признаки — пассивное сгибание или разгибание конечностей.
Глазной нистагм (пилообразные движения глаз), способствует сохранению зрительной ориентации. Нистагм имеет две фазы. При вращении сначала происходит медленное отклонение глазных яблок в сторону, противоположную вращению. Затем глаза быстро движутся обратно — по ходу вращения «догоняют организм». Медленное отклонение вызывается с рецепторов полукружных каналов, быстрый компонент связан с влиянием корковых центров (при наркозе он исчезает). Во врачебной практике направление нистагма обозначают по быстрому компоненту. Исследование нистагма (степени его выраженности, длительности) используется для оценки функционального состояния вестибулярного анализатора и дифференциальной диагностики поражения ЦНС.
Перераспределение мышечного тонуса при ходьбе и беге обеспечивает сохранение равновесия и естественной позы. Например, при сгибании одной конечности тонус мышц-разгибателей другой конечности повышается. Мезэнцефалический организм имеет нормальный тонус мышц, но у него отсутствуют произвольные движения. Однако в среднем мозге, как и в спинном, имеются нейронные цепи, в которых запрограммированы ритмические движения конечностей (локомоция); они возникают при стимуляции соответствующих центров. В нормальных условиях их деятельность запускается корой большого мозга.
Дата добавления: 2016-02-16; просмотров: 5410;