Соматические рефлексы спинного мозга
Вегетативные рефлексы, в том числе осуществляемые с помощью спинного мозга, изучаются по главам, в разделах, посвященных регуляции функций органов и систем.
Общая характеристика. Соматические рефлексы спинного мозга достаточно просты. По форме это в основном сгибательные и разгибательные рефлексы сегментарного характера. Надсегментарные рефлексы, наряду с сегментарными, осуществляются только с помощью шейного отдела.
Рефлексы спинного мозга можно объединить в две группы по следующим признакам. Во-первых, спинальные рефлексы целесообразно объединить по органам (эффекторам рефлекса): рефлексы конечностей, брюшные, яичковый и анальный. Во-вторых, по рецепторам, раздражение которых вызывает рефлекс: проприоцептивные, висцероцептивные и кожные рефлексы. Последние (кожные) являются защитными. Рефлексы, возникающие с проприорецепторов, участвуют в формировании акта ходьбы и регуляции мышечного тонуса. Висцероцептивные рефлексы возникают с интерорецепторов (рецепторов внутренних органов) и проявляются в сокращениях мышц передней брюшной стенки, грудной клетки и разгибателей спины.
Защитные рефлексы, возникающие при раздражении кожи и тела, представлены в табл. 8 и 9.
Таблица 8. Защитные брющные рефлексы, вызываемые раздражением кожи живота, выражающиеся в сокращении соответсвтующих участков мышц брюшной стенки
| Название рефлекса | Локализация рефлексогенной зоны | Сегмент спинного мозга |
| Верхний | Параллельно нижним ребрам, непосредственно под ними | Th8-Th9 |
| Средний | На уровне пупка, горизонтально | Th9-Th10 |
| Нижний | Параллельно паховой складке, рядо с ней | Th11-Th12 |
Таблица 9. Яичковый и анальный защитные рефлексы
| Название и сущность рефлекса | Локализация рефлексогенной зоны (штриховое раздражение) | Сегмент спинного мозга |
| Кремастерный (яичковый) рефлекс, выражается в сокращении m. Cremaster и поднимающая мошонки | Верхняя внутрення поверхность кожи бедра | L1-L2 |
| Анальный рефлекс, выражается в сокращении наружного сфинктера прямой кишки | Кожа вблизи заднего прохода | S2-S4 |
Рефлексы конечностей в клинической практике исследуют наиболее часто. Общепринятой классификации этих рефлексов не существует, их описывают без взаимной связи. Если за объединяющий признак рефлексов конечностей принять характер ответной реакции, то все их можно объединить в четыре группы.
1. Сгибательные рефлексы конечностей (фазные и тонические).
Фазные рефлексы — это однократное сгибание конечности при однократном раздражении кожи или мышечных рецепторов. Рефлексы, возникающие с рецепторов кожи, являются полисинаптическими, они имеют защитное значение. Например, погружение в слабый раствор серной кислоты кончика лапки спинальной лягушки, подвешенной на крючке, или щипок кожи конечности пинцетом вызывают отдергивание конечности вследствие сгибания ее в коленном суставе, а при более сильном раздражении — и в тазобедренном суставе (сгибание конечности — избавление от раздражителя). Сгибательные (как и разгибательные) рефлексы, возникающие с мышечных рецепторов, являются моносинаптическими. Фазные проприоцептивные рефлексы участвуют в формировании акта ходьбы. По степени выраженности фазных сгибательных и разгибательных рефлексов определяют состояние возбудимости ЦНС и возможные ее нарушения. Поэтому в клинической практике исследуют несколько сгибательных фазных рефлексов. Ахиллов (пяточный) рефлекс— проприоцептивный, выражается в подошвенном сгибании стопы в результате сокращения трехглавой мышцы голени, возникает при ударе молоточком по ахиллову сухожилию; рефлекторная дуга замыкается на уровне крестцовых сегментов S1 —S2. Подошвенный рефлекс — кожный, выражается в сгибании стопы и пальцев при штриховом раздражении подошвы; рефлекторная дуга замыкается также на уровне S1-S2.
Наиболее часто в клинической практике исследуют локтевой рефлекс — проприоцептивный, выражается в сгибании руки в локтевом суставе, возникает при ударе молоточком по сухожилию m. biceps brachii (при вызове рефлекса рука должна быть слегка согнута в локтевом суставе); рефлекторная дуга замыкается в шейных сегментах спинного мозга (С5—С6). При возникновении фазных сгибательных рефлексов одновременно с возбуждением мотонейронов мышц-сгибателей происходит торможение мотонейронов мышц-разгибателей этой же конечности — иначе сгибание будет невозможным.
Тонические сгибательные рефлексы конечностей участвуют в поддержании позы. Они возникают при длительном растяжении мышц и возбуждении мышечных рецепторов. Тоническое сокращение скелетных мышц является также фоновым для всех двигательных актов, осуществляемых с помощью фазных сокращений мышц. Изменение тонуса мышц конечностей свидетельствует о нарушениях в ЦНС.
2. Разгибательные рефлексы конечностей — моносинаптические, возникают с проприорецепторов мышц-разгибателей.
Фазные разгибательные рефлексы, в отличие от фазных сгибательных рефлексов, возникают в ответ на однократное раздражение только мышечных рецепторов, например при ударе по сухожилию четырехглавой мышцы ниже надколенника. При этом наблюдается коленный разгибательный рефлекс вследствие сокращения четырехглавой мышцы (мотонейроны мышц-сгибателей во время разгибательного рефлекса тормозятся с помощью постсинаптического реципрокного торможения посредством вставочных тормозных клеток Реншоу). Рефлекторная дуга коленного разгибательного рефлекса замыкается в поясничных сегментах (L2—L4). Фазные разгибательные рефлексы, как и сгибательные, участвуют в формировании акта ходьбы (перемещение тела в пространстве).
Тонические разгибательные рефлексы — это длительное сокращение мышц-разгибателей при длительном их растяжении и возбуждении мышечных рецепторов. В положении стоя тоническое сокращение мышц-разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тоническое сокращение мышц спины удерживает туловище в вертикальном положении, обеспечивая осанку человека. Тонические рефлексы на растяжение мышц (сгибателей и разгибателей) называют также миотатическими.
Таким образом, главная роль тонических рефлексов — поддержание позы организма.
3. Ритмические рефлексы выражаются в многократном повторном сгибании и разгибании конечностей. Примером является рефлекс потирания: после смазывания раствором кислоты кожи бедра спинальная лягушка (лягушка, у которой удален головной мозг) многократно потирает этот участок — пытается освободиться от раздражителя. Слабое раздражение кожи боковой поверхности тела у собаки также вызывает ритмический рефлекс — почесывание этого участка задней конечностью. У спинального животного наблюдается также шагательный рефлекс.
Шагательный рефлекс вызывается однократным раздражением кожи задней конечности, которое, в свою очередь, вызывает сгибание последней и сопровождается дополнительным разгибанием противоположной задней конечности — перекрестный разгибательный рефлекс. Затем согнутая конечность разгибается и опускается вниз, а разогнутая — сгибается и поднимается вверх, и так далее. Конечности не касаются опоры (собака зафиксирована в станке с помощью лямок), и без дополнительного раздражения они продолжают ритмично поочередно сгибаться и разгибаться — «шагать» в воздухе (рис. 45).
Шагательный рефлекс (локомоция) может продолжаться часами, так как устранено тормозное влияние коры большого мозга. При неповрежденных сегментах, иннервирующих передние конечности (С5 - Тh2), и более длительном раздражении задней конечности в рефлекс вовлекаются также и передние конечности. Это наблюдение показывает наличие межсегментарной связи на уровне спинного мозга, играющей важную роль в ритмических движениях конечностей.
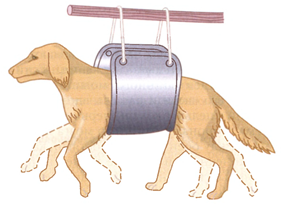
Рис. 45. Диагональные шагательные движения, выполняемые спинальным животным (ритмический рефлекс) (Гайтон, 2008)
Поочередное сокращение и расслабление мышц-сгибателей и мышц-разгибателей осуществляется в результате взаимодействия процессов возбуждения и торможения в соответствующих центрах спинного мозга под влиянием импульсов, поступающих в мозг от проприорецепторов, что определяется их особым расположением относительно скелетной мышцы. Мышечные веретена (мышечные рецепторы, они первичные) расположены параллельно скелетной мышце — своими концами они крепятся к соединительнотканной оболочке (перимизию) пучка экстрафузальных мышечных волокон с помощью напоминающих сухожилия полосок соединительной ткани длиной 0,5—1 мм. В связи с этим, когда мышца расслабляется (удлиняется), растягиваются и мышечные рецепторы, что и ведет к их возбуждению (см. рис. 6.11, A). Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон (fusus — веретено), окруженных соединительнотканной капсулой и окончаниями афферентных волокон. Длина интрафузальных мышечных волокон составляет 4—7 мм, толщина — 15—30 мкм. Импульсы от мышечных рецепторов возбуждают нейроны своего центра и тормозят нейроны центра-антагониста. Ближе к концам мышечного веретена подходят двигательные нервные окончания, являющиеся аксонами γ-мотонейронов спинного мозга. Их импульсация вызывает сокращение интрафузальных мышечных волокон (мышечные волокна рецептора), что ведет к возникновению РП, инициирующего посредством своего электрического поля возбуждение афферентного нервного волокна.
Сухожильные рецепторы (рецепторы Гольджи) крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение при натяжении сухожилия (рис. 46). Поэтому сухожильные рецепторы посылают в мозг информацию о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы — что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее).
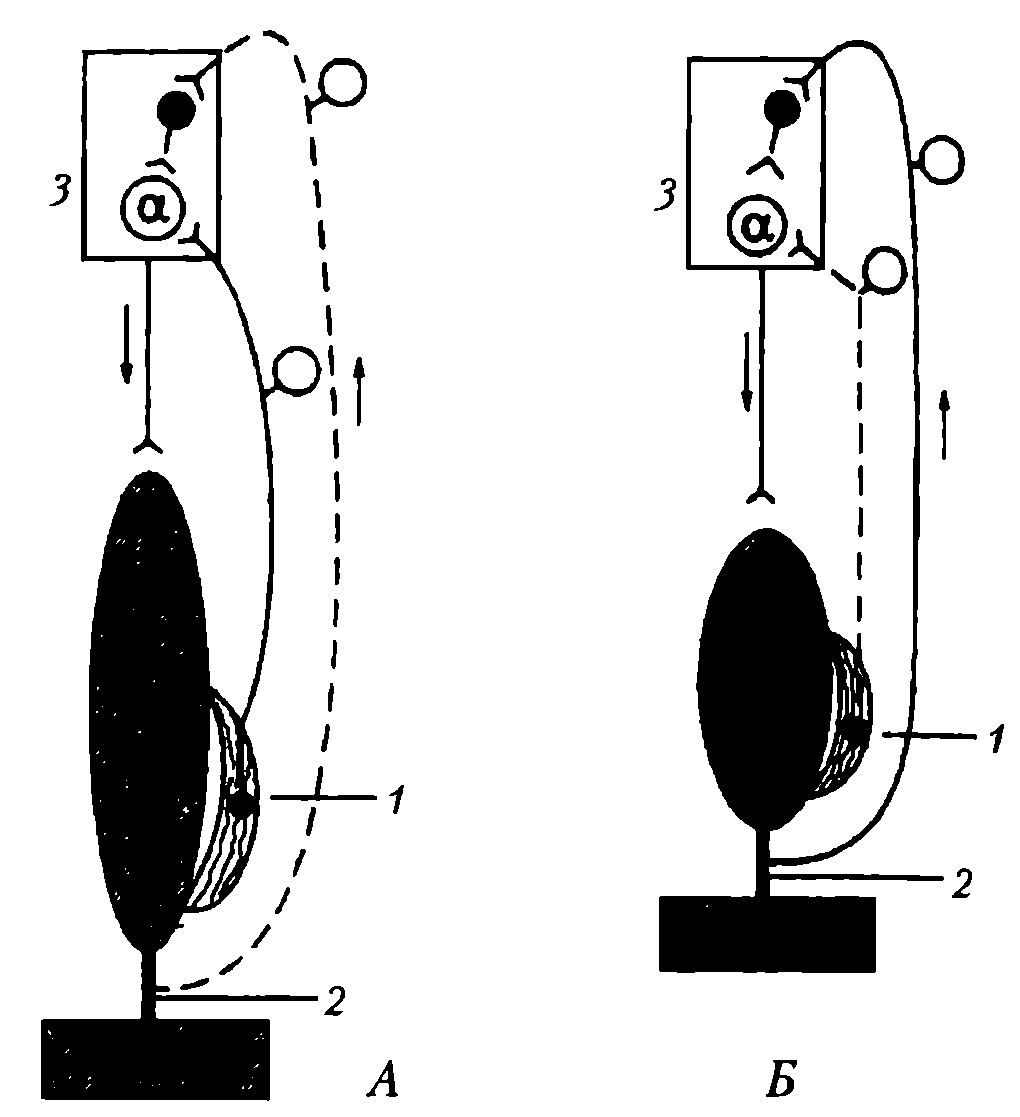
Рис. 46. Механизм возбуждения и торможения α-мотонейронов спинного мозга во время сокращения и расслабления скелетной мышцы: А – мышца растянута и расслаблена, возбуждаются мышечные рецепторы (1); Б – мышца сокращена (напряжена) и укорочена, возбуждаются сухожильные рецепторы (2); 1 – мышечные рецепторы (мышечные веретена); 2 – сухожилия мышцы и их рецепторы (рецепторы Гольджи); 3 – полусегмент спинного мозга. Афферентная импульсация: сплошная линия – выражена, пунктирная линия - отсутствует
Сухожильные рецепторы заключены в соединительнотканную капсулу и локализуются в сухожилиях скелетных мышц вблизи от сухожильно-мышечного соединения. Это тоже первичные рецепторы, они представляют собой безмиелиновые окончания толстого миелинового афферентного волокна — подойдя к капсуле рецепторов Гольджи, это волокно теряет миелиновую оболочку и делится на несколько окончаний, лишенных миелина.
Механизм шагательного рефлекса. Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим α-мотонейронам спинного мозга и возбуждают их. Затем α-мотонейроны посылают импульсы к этой же скелетной мышце, что ведет к ее сокращению. В результате сокращения мышцы возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), начинают возбуждаться сухожильные рецепторы, поскольку сухожилие натягивается.
Импульсы от сухожильных рецепторов поступают тоже, в первую очередь, в свой центр в спинном мозге, но к тормозным клеткам. Возбуждение этих клеток вызывает торможение α-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление (удлинение) ведет снова к возбуждению мышечных веретен и α-мотонейронов — мышца вновь сокращается. Вследствие ее сокращения снова возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что ведет к очередному расслаблению скелетной мышцы, и так далее. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю, при этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу, что наблюдается при ходьбе и беге или любом повторяющемся действии.
Таким образом, и мышцы-сгибатели, и мышцы-разгибатели рефлекторно сокращаются только с помощью мышечных рецепторов, а рефлекторно расслабляются только с помощью сухожильных рецепторов. Во время ходьбы в естественных условиях мышцы-сгибатели и мышцы-разгибатели одной и той же конечности должны сокращаться и расслабляться последовательно друг за другом, что достигается с помощью торможения центра-антагониста при возбуждении центра-агониста. Причем если на одной ноге сокращены сгибатели, на другой ноге сокращаются разгибатели, иначе нарушится поза. Это обеспечивается поступлением афферентных импульсов от мышечных и сухожильных рецепторов и поочередным возбуждением и торможением центров-сгибателей и центров-разгибателей — перекрестный разгибательный рефлекс. Шагательные координированные движения возможны в отсутствие обратной афферентации от проприорецепторов. Они осуществляются с помощью межсегментарных связей на уровне спинного мозга и генератора локомоции каждой конечности. О наличии межсегментарных связей на уровне спинного мозга свидетельствует также факт вовлечения в шагательный рефлекс всех четырех конечностей при достаточно длительном и сильном раздражении одной конечности. Однако у человека после разрыва спинного мозга локомоция не восстанавливается, но эти механизмы включаются вышележащими отделами ЦНС.
4.Рефлексы позы спинального организма (шейные рефлексы) (рис. 47).
Они заключаются в перераспределении тонуса мышц конечностей при изменении положения шеи относительно туловища.
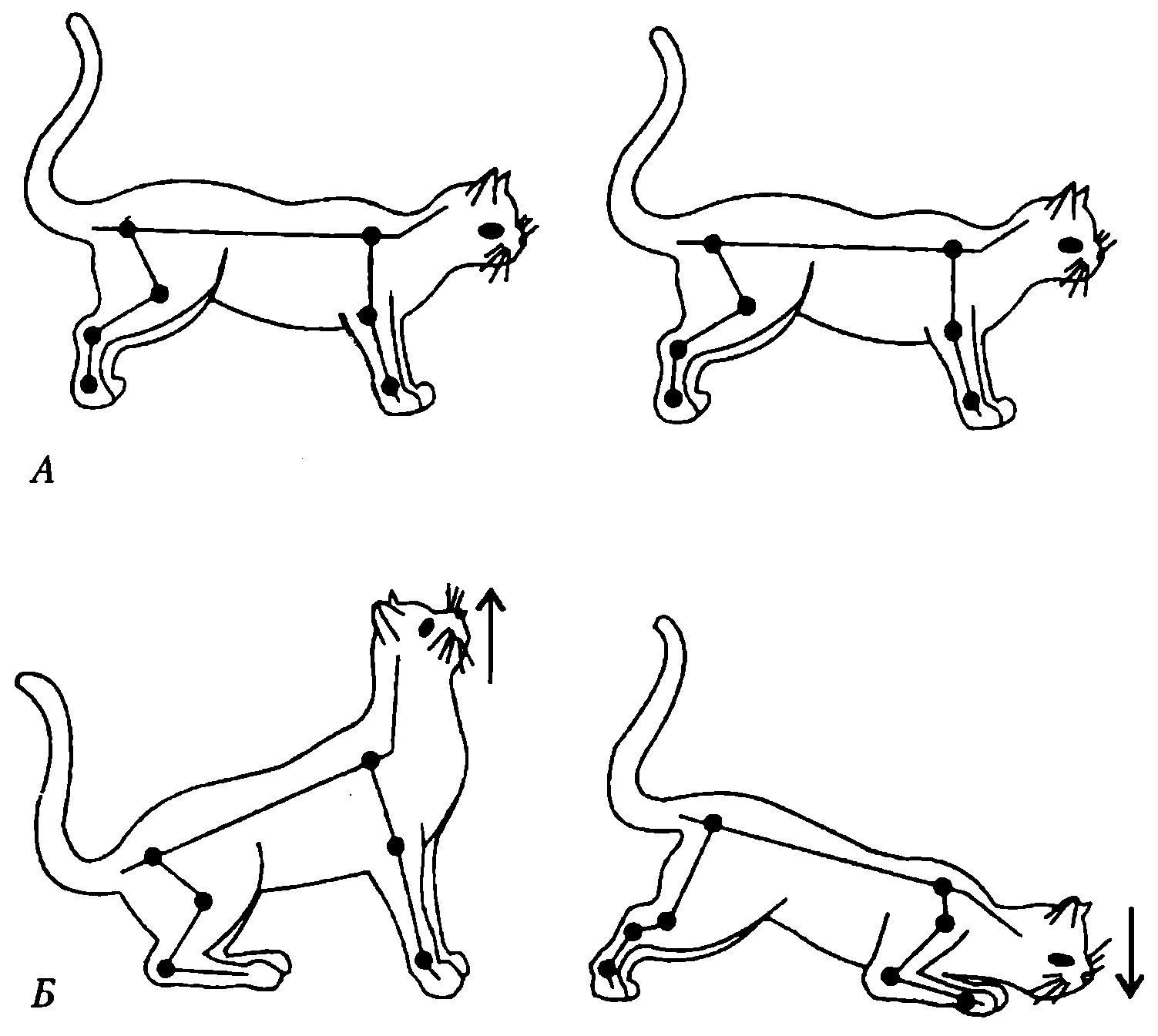
Рис. 47. Позные шейные рефлексы у кошки с удаленным вестибулярным аппаратом: А – до изменения положения головы; Б – при пассивных (↑) и опускании (↓) головы
Они осуществляются с участием различных отделов ЦНС. На уровне спинного мозга замыкаются шейные позные рефлексы, наличие которых установил голландский физиолог Р. Магнус (1924) в опытах на кошке. Имеется две группы этих рефлексов.
Первая группа шейных позных рефлексов возникает при наклоне головы вперед или назад (рис. 47).
При наклоне головы вниз (кпереди) увеличивается тонус мышц-сгибателей передних конечностей и тонус мышц-разгибателей задних конечностей, в результате чего передние конечности сгибаются, а задние — разгибаются. При наклоне головы вверх (кзади) возникают противоположные реакции: передние конечности разгибаются вследствие увеличения тонуса их мышц-разгибателей, а задние конечности сгибаются вследствие повышения тонуса их мышц-сгибателей. Эти рефлексы возникают с проприорецепторов мышц шеи и фасций, покрывающих шейный отдел позвоночника. В условиях естественного поведения животных они увеличивают шансы достать пищу, находящуюся выше или ниже головы, — это автоматизированная приспособительная реакция, рефлексы являются надсегментарными. Шейные рефлексы позы верхних конечностей у человека, по-видимому, утрачены, а рефлексы нижних конечностей выражаются не в сгибании или разгибании, а в перераспределении мышечного тонуса, обеспечивающего сохранение естественной позы.
Вторая группа шейных позных рефлексов (тонических) возникает с шейных проприорецепторов при поворотах или наклонах головы вправо или влево (рис. 48).
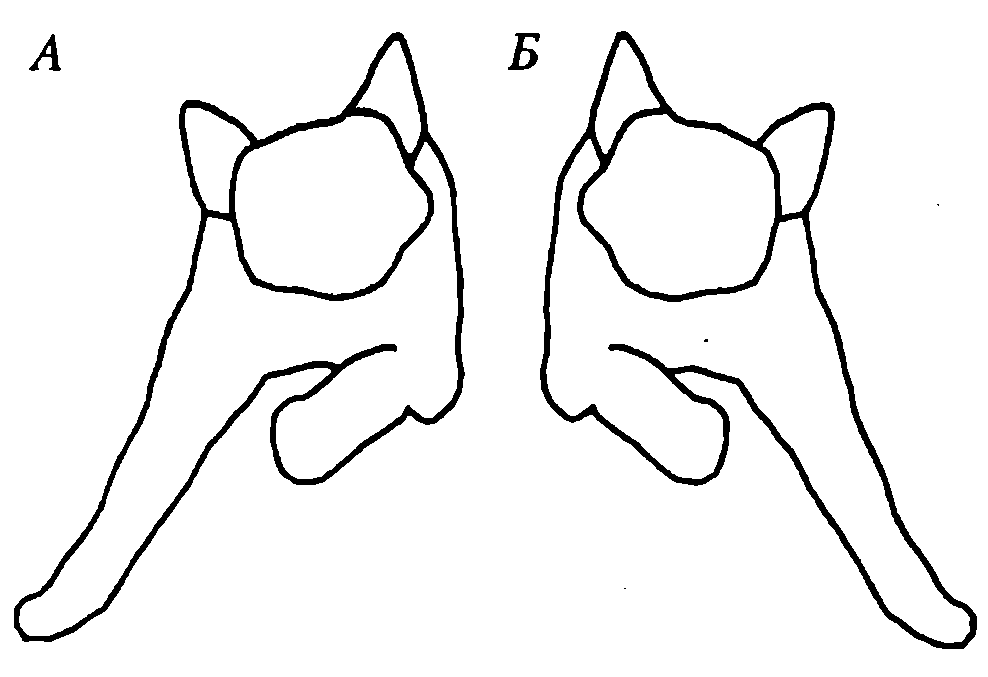
Рис. 48. Изменеие тонуса мышц конечностей при наклоне головы вправо (А) и влево (Б) (шейные тонические рефлексы)
Рефлекс выражается в повышении тонуса мышц-разгибателей обеих конечностей на стороне, в которую повернута (наклонена) голова, и в повышении тонуса мышц-сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие изменения положения центра тяжести после поворота (наклона) головы. Центр тяжести смещается в сторону поворота головы — именно на этой стороне повышается тонус мышц-разгибателей обеих конечностей, что повышает устойчивость организма. Подобная ситуация наблюдается и у человека.
Дата добавления: 2016-02-16; просмотров: 3325;