И МЕХАНИЗМ ЕЕ СОКРАЩЕНИЯ
Структурной и функциональной единицей скелетной мышцы является мышечное волокно, представляющее собой сильно вытянутую многоядерную клетку. Длина мышечного волокна зависит от размеров мышцы и составляет от нескольких милли-
|
метров до нескольких сантиметров. Толщина волокна вариабельна (10-100 мкм). Скелетная мышца - это система, преобразующая химическую энергию в механическую работу и тепло.
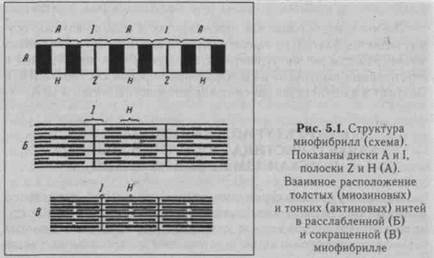
Специфическими элементами мышечного волокна являются специализированный сократительный аппарат - миофибриллы, системы продольных трубочек - саркоплазматическая сеть (ретикулум) и система поперечных трубочек - Т-система, представляющая собой впячивания поверхностной мембраны мышечного волокна внутрь его. Миофибрилла состоит из нескольких саркомеров, являющихся функциональной единицей сократительного аппарата мышечного волокна. Саркомеры отделяются друг от друга 2-пластинками.
Скелетная мышца имеет поперечную исчерченность, которая обусловлена особым расположением сократительных миофиб-рилл - белков актина и миозина: их концы не полностью перекрывают друг друга, что и определяет исчерченность (рис. 5.1). Акти-новые филаменты представлены двойной нитью, закрученной в двойную спираль; они одним концом прикреплены к 2-пластинке, другим концом взаимодействуют с нитями миозина при сокращении мышцы. Нити миозина располагаются в середине саркомера и своими концами взаимодействуют с нитями актина. В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина, к молекуле которого прикреплена молекула другого белка - тропонина. В состоянии покоя нити актина и миозина незначительно перекрывают друг друга таким образом, что общая длина саркомера составляет около 2,5 мкм.
На боковых сторонах миозиновой нити имеются выступы, получившие название поперечных мостиков, состоящих из головки и шейки. Головка приобретает выраженную АТФ-азную активность при контакте с актином.
При сокращении происходит укорочение саркомера вследствие активного перемещения тонких актиновых нитей относительно толстых миозиновых.
Механизм мышечного сокращения. Мышца сокращается в естественных условиях только при поступлении к ней нервных импульсов. Нервное влияние на мышечное волокно передается с помощью нервно-мышечного синапса. Медиатором в нервно-мышечном синапсе является ацетилхолин. На один ПД из пресинаптиче-ского окончания нервно-мышечного синапса выделяется 200-300 квантов медиатора.
В состоянии покоя мышцы, т.е. в промежутках между передачей нервного импульса, происходит спонтанное выделение 1-2 квантов медиатора в синаптическую щель в среднем 1 раз в секунду. При этом на постсинаптической мембране формируется деполяризация с амплитудой 0,12-0,24 мВ. Такие потенциалы получили название миниатюрные потенциалы концевой пластинки. Они, вероятно, поддерживают высокую возбудимость синапсов в условиях функционального покоя нервных центров. Кроме эк-зоцитоза медиатора существует постоянная неквантовая утечка молекул медиатора в синаптическую щель. Предполагают, что неквантовая секреция играет трофическую роль.
Пришедший по нервному волокну импульс (ПД) обеспечивает выделение в синаптическую щель ацетилхолина, который на пост-• синаптической мембране (концевой пластинке мышечного волокна) вызывает возникновение потенциала концевой пластинки (ПКП) -возбуждающего постсинаптического потенциала (ВПСП), амплитуда которого составляет 30-40 мВ. ПКП - это локальный потенциал, который, достигнув критической величины, обеспечивает возникновение ПД в мышечном волокне. ПД распространяется по мышечному волокну и Т-системе в глубь волокна, что обеспечивает выделение ионов Са2+ из саркоплазматического ретикулума. При взаимодействии ионов Са2+ с тропонином происходят смещение тропомиозина и освобождение активных центров на актиновых нитях.
Затем происходит присоединение головки поперечного мостика миозина к актиновой нити. При этом головка миозина приобретает АТФ-азную активность, что обеспечивает гидролиз АТФ и освобождение энергии, обеспечивающей поворот головки миозина вокруг своей оси (гребковое действие), что в свою очередь приво-
дит к скольжению нитей актина и миозина относительно друг друга и укорочению саркомера и общей длины мышцы. Миозиновая головка имеет несколько активных центров, которые последовательно взаимодействуют с соответствующими центрами на актиновой нити. В каждый конкретный момент в процессе развития сокращения одни головки поперечных мостиков находятся в соединении с актиновой нитью, другие свободны, т. е. существует последовательность их взаимодействия с актиновой нитью, что обеспечивает плавность процесса сокращения.
Повторное присоединение миозиновой головки к новому центру на актиновой нити вновь приводит к повороту головки, который обеспечивается запасенной в ней энергией. В каждом цикле соединения и разъединения головки миозина с актином расщепляется одна молекула АТФ на каждый мостик. Быстрота поворота определяется скоростью расщепления АТФ.
Для расслабления мышцы в первую очередь необходимо понижение концентрации ионов Са2+ в области сократительных элементов мышечного волокна. Саркоплазматическая сеть имеет кальциевый насос, который активно возвращает кальций в цистерны. Активация кальциевого насоса осуществляется неорганическим фосфатом, который образуется при гидролизе АТФ, а энергообеспечение работы кальциевого насоса также осуществляется за сче* энергии, образующейся при гидролизе АТФ.
Источником энергии для восстановления израсходованной АТФ являются белки, жиры и углеводы пищи, которые подвергаются расщеплению в желудочно-кишечном тракте и в виде мономеров поступают в кровь и лимфу.
В организме в результате биохимических превращений образуется АТФ или синтезируются крупномолекулярные вещества. АТФ - донор свободной энергии в клетках. В клетках АТФ используется в течение одной минуты после ее образования, скорость оборота АТФ очень велика. Стабильность концентрации АТФ в клетке поддерживается рядом механизмов, одним из которых является образование креатинфосфата (КФ). Когда количество АТФ превышает определенный уровень, часть ее энергии используется для синтеза КФ, количество которого при этом возрастает. При повышении же распада АТФ в условиях активации энергетического обмена КФ используется для ресинтеза АТФ с помощью окисления и фосфорилирования.
Существуют и бескислородные (анаэробные) пути преобразования энергии, в которых могут быть использованы только углеводы (анаэробный гликолиз); такие способы реализуются при недостаточном поступлении кислорода в организм, ткани и клетки. При
полном прекращении дыхания и расходовании резервов кислорода эти процессы могут обеспечить потребность в энергии еще в течение двух минут.
Запас АТФ в скелетных мышцах обеспечивает всего лишь 10 одиночных сокращений. При максимальном мышечном сокращении имеющихся в тканях запасов АТФ достаточно лишь на одну секунду. Энергия КФ, концентрация которого в три-восемь раз больше, чем АТФ, может поддержать такое сокращение в течение еще нескольких секунд. При максимальном сокращении на протяжении нескольких секунд абсолютно необходим анаэробный гликолиз, в котором используются запасы гликогена. Ресинтез гликогена из образующейся при этом молочной кислоты возможен, однако, лишь в аэробных условиях.
Аэробное окисление глюкозы и жирных кислот в цикле Кребса, совершаемое в митохондриях, - это наиболее типичный способ энергообеспечения скелетных мышц. Запасы свободной глюкозы, гликогена и жиров в мышцах достаточно велики. Однако при длительной работе в организме накапливаются недоокисленные продукты (молочная кислота и др.). Создается кислородная задолженность. Такой долг погашается после работы за счет компенсаторной мобилизации кровообращения и дыхания (тахикардия, повышение кровяного давления, одышка). Если же работа, несмотря на наличие кислородного долга, продолжается, наступает выраженное состояние утомления, которое иногда прекращается за счет мобилизации дополнительных резервов кровообращения и дыхания («второе дыхание» спортсменов).
Таким образом, энергия АТФ в скелетной мышце используется для трех процессов: 1) работы № / К-насоса, обеспечивающего поддержание постоянства градиента концентрации этих ионов по обе стороны мембраны; 2) процесса скольжения актиновых и миозино-вых нитей, приводящих к укорочению миофибрилл (комплекс актин - миозин становится стабильным только при трупном окоченении, когда концентрация АТФ в мышце падает ниже некоторой критической величины)* 3) работы Са-насоса, активируемого при расслаблении волокна.
Дата добавления: 2016-04-11; просмотров: 754;