Модельные мембранные системы. Монослой на границе раздела фаз. Бислойные мембраны. Протеолипосомы
Мультиламеллярные липосомы - при энергичном встряхивании образуют замкнутые сферические частицы. Этот процесс энергетически выгоден, так как в этом случае достигается минимум площади контакта гидрофобных поверхностей с водой. При простых механических воздействиях образуются в основном многослойные частицы (диаметром в несколько микрометров) с концентрической организацией слоев - мультиламеллярные липосомы или везикулы (мешочки). В этих частицах липидные бислои отделяют внутреннюю водную фазу от внешнего раствора. Такая организация по-зволяет использовать мультиламеллярные липосомы для исследования барьерных свойств липидного. Мультиламеллярные липосомы осмотически активны: они изменяются в объеме при изменении осмотических свойств внешней среды.
Моноламеллярные липосомы. Более энергичное воздействие на грубые фосфолипидные дисперсии, например с помощью ультразвукового излучения, вызывает образование моноламеллярных (однослойных) везикул. (d=20-40 нм). Моноламеллярные липосомы используют в исследованиях биологического и медико-биологического характера. Тем не менее, малый внутренний объем и осмотическая неактивность липосом существенно сужают круг этих исследований. В последнее время разработаны методики получения больших (100 нм в диаметре и более) моноламеллярных липосом, которые лишены этих недостатков.
Протеолипосомы. Многие мембранные белки и мелкие фрагменты биологических мембран сравнительно легко могут быть включены в состав искусственных везикулярных мембран. Такие комбинированные системы называются протеолипосомами. Эффективность встраивания большинства белков компонентов в искусственные мембранные системы резко зависит от липидного состава мембран, pH, солевого состава, температуры и т. д. Как правило, эффективность встраивания белков или липопротеиновых фрагментов мембраны возрастает в присутствии небольших количеств детергентов. Полагают, что мелкие молекулы детергентов заполняют дефекты на границе белковой молекулы в мембране, экранируя гидрофобные участки мембраны от воды.
Плоские бислоиные липидные мембраны. Липиды, спонтанно образующие ла-меллярные слои, обычно способны формировать бислоиные структуры (БЛМ или «черные» пленки) на небольших отверстиях в тонких гидрофобных материалах. Относительная простота получения БЛМ, широкий спектр применения разнообразных электроизмерительных методов исследования, возможность изменять в широких пределах липидный состав БЛМ и состав омывающих растворов, включать в БЛМ разнообразные модификаторы барьерных свойств мембран, функционально активные элементы биологических мембран - все это быстро обеспечило этим искусственным мембранным системам центральное место в современной экспериментальной мембранологии.
Общая схема ячейки для формирования БЛМ и несколько последовательных этапов формирования бислойной структуры:
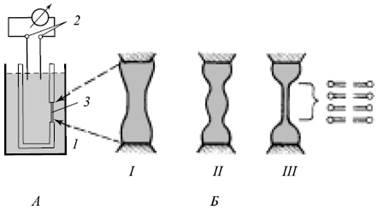
Образование БЛМ: А – ячейка для получения БЛМ и изучения ее электрических св-в
Б - этапы формирования БЛМ (Ш = БЛМ)
1 - тефлоновый стакан
2 - электроды
3 - отверстие в стенке стакана
Процесс образования БЛМ начинается с нанесения небольшого количества подходящего липида, растворенного обычно в жидких углеводородах (гексан, октан, декан), на отверстие в тефлоновом стаканчике. Далее эта толстая пленка самопроизвольно утоньшается вплоть до образования бислойной структуры. Главными силами, определяющими поведение пленки, являются межфазное (поверхностное) натяжение и ван-дер-ваальсовы силы притяжения водных фаз по обе стороны пленки, сжимающие пленку в поперечном направлении, - расклинивающее давление.
По своим электрическим характеристикам бислойные липидные пленки, приготовленные из природных липидов, близки к биологическим мембранам. Немодифицированные искусственные БЛМ отличаются низкой проводимостью. Они лишены метаболической активности и не обладают столь высокой селективностью, как биологические мембраны. Однако многие из этих свойств могут быть приданы искусственным мембранам путем введения в их состав разнообразных мембранноактивных соединений и отдельных функционально важных элементов биологических мембран. В присутствии всех этих модификаторов проводимость бислойных мембран обычно возрастает. Все это свидетельствует о том, что бислойная липидная структура выполняет роль естественной матрицы для многих молекулярных компонентов биологических мембран и что липидный бислой в той или иной мере свойственен природным мембранам.
Основные закономерности транспорта ионов через мембраны изучены в опытах с различными моделями, из которых самой близкой к биомембранам оказалась бислойная липидная мембрана.
Затраты энергии, необходимые для проникновения иона в неполярную фазу, можно оценить по формуле Борна:
Энергия заряда иона раиусом r в среде с диэлектрической проницаемостью эпсилон=z^2*e^2/ (8пи*r*эпсилон*эпсилон0), где эпсилон0-абсолютная диэлэлектрическая проницаемость вакуума, т е
энергия, затрачиваемая на перемещение иона из воды в мембрану, зависит от его радиуса r и диэлектрических проницаемостей воды ев и мембраны ем.
В системе СГС эта формула имеет наиболее простую и наглядную запись:
Энегрия заряда иона раиусом r в среде с диэлектрической проницаемостью эпсилон=z^2*e^2/(2r) (1/ эпсилонм - 1/ эпсилонB )
где z- валентность иона, е - элементарный заряд.
Рассчитанные по данному уравнению значения свободной энергии для перехода K + и K + из воды в неполярный растворитель с эпсилонм = 2 составляют большую величину (250 - 350 кДж/моль). Именно это создает барьер, препятствующий прохождению ионов щелочных металлов через гидрофобную часть липидного бислоя в негидратированной форме.
Энергия перехода иона в мембрану снижается с увеличением радиуса иона. Поэтому крупные органические ионы проникают через БЛМ легче, чем катионы щелочных металлов. Для таких крупных липофильных ионов, как дипикриламин и тетрафенилбор, возможно прямое прохождение через мембрану. При изменении градиента концентрации в случае одного проникающего иона разность электрических потенциалов на БЛМ изменяется в соответствии с уравнением Нернста. Этим методом было продемонстрировано пассивное движение проникающих ионов через мембраны митохондий, субмитохондриальных частиц, бактериальных хроматофоров при энергозависимой генерации мембранного потенциала.
Как видно из, величина энергетического барьера в мембране уменьшается, а следовательно, проницаемость мембраны для иона возрастает не только при увеличении его радиуса, но и при приближении значений эпсилонм к эпсилонв. На этих физических принципах и основан перенос ионов ионофорами. Ионофоры могут образовывать с ионом комплекс большого размера – переносчики - либо формировать пору в мембране, заполненную водой, - каналы. Это механизмы переноса - с участием переносчиков и через ионные каналы - изучены наиболее подробно в опытах с БЛМ. (см 5.3, правда не очень много там)
Монослои.
Основные мембранообразующие липиды представляют собой соединения с идеальным сочетанием гидрофобных и гидрофильных свойств. Они сравнительно плохо растворимы в воде в мономерном виде, а стремление их полярных головок максимально контактировать с водой придает им уникальные способности образовывать многообразные сравнительно устойчивые структуры при агрегации этих молекул.
Нерастворимые монослои на границе раздела вода-воздух. Большинство мембранообразующих липидов при нанесении их на поверхность воды практически не растворяется в объемной фазе, а быстро растекается по поверхности, образуя мономолекулярный слой. При сжатии монослоя начинают проявляться межмолекулярные взаимодействия, нарушающие состояние идеального двумерного газа.
Дальнейшее возрастание давления приводит к формированию так называемой растянутой жидкой пленки. Молекулы в этом состоянии обладают всеми степенями свободы, как и в обычной жидкости, однако их полярные головки фиксированы на поверхности раздела фаз. Углеводородные цепи молекул в этом состоянии склонны к агрегации и взаимопроникновению. При дальнейшем возрастании давления в монослое будет реализовываться наиболее плотная упаковка молекул, т. е. их вертикальная ориентация. Вначале подвижность углеводородных цепей будет выше, чем у полярных головок, так как последние сильно гидратированы. В этом состоянии сохраняется возможность движения молекул в плоскости поверхности (латеральная подвижность) и его можно рассматривать как конденсированную жидкую пленку:
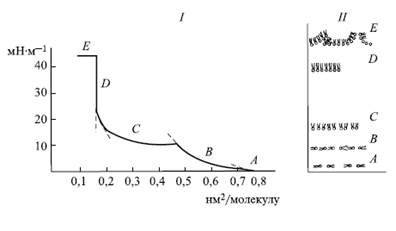
Кривая (изотерма) площадь-давление для монослоя длинноцепочных липидов (1) и схема расположения липидных молекул на границе раздела фаз вода-воздух (2):
А - состояние двумерного газа,
В - растянутая жидкая пленка,
С - конденсированная жидкая пленка,
Б - твердоконденсированное состояние,
Е - состояние коллапса
уплотнение слоя при возрастании давления может произойти лишь за счет разрушения гидратных оболочек полярных головок, после чего слой становится практически несжимаемым и находится в твердоконденсированном состоянии, где площадь поперечного сечения молекул не зависит от длины углеводородной цепи. После превышения некоторого предельного давления монослой начинает разрушаться -состояние коллапса.
Поверхностные монослои широко используют в качестве модельных мембранных систем. С их помощью изучают подвижность и типы упаковки молекулярных компонентов в мембранах, межмолекулярные взаимодействия в мембранах, механические свойства мембран; исследуют кинетику и механизмы ферментативных процессов, протекающих на границе раздела фаз; изучают процессы переноса ионов и электронов через границу раздела фаз, инжекцию заряда в липидный слой (диэлектрик). Однако этот метод имеет ряд ограничений, в значительной степени обусловленных тем, что монослой - это лишь половина липидного слоя мембран, обращенного в газовую фазу. Последнего ограничения удается избежать при использовании в качестве мембраны мономолекулярного слоя, образующегося на границе двух несмешивающихся жидкостей (углеводород-вода). Более адекватные модели, представляющие собой липидные бислои, удается получить в виде полимолекулярных структур, которые образуются липидами в объеме водной фазы.
| <== предыдущая лекция | | | следующая лекция ==> |
| | | Изменение энтропии в открытых системах. Постулат Пригожина. Термодинамические условия осуществления стационарного состояния. Примеры |
Дата добавления: 2016-04-11; просмотров: 5077;