Структурная организация и функционирование фотосинтетических мембран. Фотосинтетическая единица. Два типа пигментных систем и две световые реакции
Важнейшая особенность топографической организации первичных процессов фотосинтеза состоит в том, что отдельные переносчики ЭТЦ в хлоропластах объединены в единые электронно-транспортные макромолекулярные комплексы, локализованные в фотосинтетических мембранах. Наибольшее значение имеют следующие из них: светособирающий пигмент-белковый комплекс (ССПБК), от которого энергия возбуждения направляется на РЦ; комплексы фотосистем ФС II (ПБК II) и ФС I (ПБК I), включающие помимо светособирающего, или антенного, хлорофилла a фотохимически активные пигменты Pб80 и -700 соответственно, а также непосредственные доноры и акцепторы электронов для РЦ фотосистем; цитохромный комплекс b 6—f, который состоит из пяти белковых субъединиц и включает цитохром f, два цитохрома 6 , а также железосерный белок Риске. Цитохромный комплекс ответствен за последовательный перенос электрона между фотосистемами и циклический транспорт электронов вокруг ФС I. АТФазный сопрягающий комплекс CF 0 — CF 1 за счёт протонного градиента на мембране синтезирует АТФ
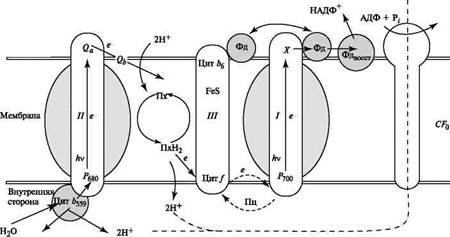 |
Рисунок 1
На схеме виден асимметричный характер поперечного расположения донорных и акцепторных компонентов ФС I и ФС II в мембране тилакоида. В последние годы стало ясно (Дж. Андерсон, Дж. Барбер), что и латеральное расположение комплексов в плоскости мембраны носит явно вы-раженный гетерогенный характер (ФС II – на мембранах, обращённых друг к другу, ФС I – на внешних мембранах гран и на мембранах стромы)
Проходя через ЭТЦ, электрон на своей пути пересекает мембрану, проходя через комплекс b 6 f от ее внутренней стороны, где расположена донорная часть ФС II, на наружную сторону к акцепторному участку ФС I. Взаимодействие между макромолекулярными комплексами ФС I, b 6 f и ФС II осуществляется при посредстве подвижных переносчиков - пластоцианина Пц и пластохинона Пх, причем молекулы последнего образуют пул, содержащий не менее 10 молекул Пх на один Пц. Другой подвижный переносчик - ферредоксин - акцептирует электроны у ФС I, осуществляя связь ППФ с темновыми процессами.
Пигментные системы, входящие в состав ФС I и ФС II, отличаются по своим спектральным свойствам, а хлорофилл ФС I поглощает в более длинноволновой области по сравнению с хлорофиллом ФС II. Это позволяет селективно возбуждать фотосистемы: отдельно ФС I (λ > 680 нм) и вместе ФС I+ФС II (λ < 680 нм).
Комплекс светособирающих пигментных молекул, передающих энергию на один фотоактивный центр (ФС I и ФС II), получил название фотосинтетической единицы (ФСЕ)
|
Основной причиной конформационных изменений в мембранах тилакоидов и перераспределения энергии возбуждения между ФС II и ФС I является изменение заряда ССПБК. Даже в отсутствие фосфорилирования светособирающего комплекса, но при низкой концентрации положительных ионов, отрицательные заряды, находящиеся на поверхности мембранных белков, создают высокий поверхностный электрический потенциал на тилакоидных мембранах и препятствуют слипанию тилакоидов. В этих условиях ФС I и ССПБК равномерно распределены в плоскости мембраны, благодаря чему возможна эффективная миграция энергии от ФС II к ФС I. Фосфорилирование ССПБК приводит к появлению на нем дополнительного отрицательного заряда, что вызывает переход ССПБК в стромальный участок мембраны и возрастание переноса энергии от него к ФС I. Фофорилирование ССПБК осуществляется с помощью протеинкиназы, находящейся в гранальных участках мембраны.
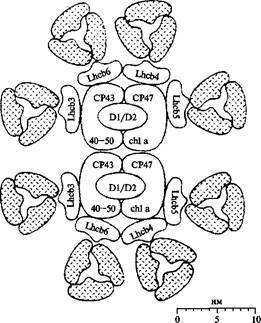
Комплекс ФСII
Рисунок 2
Внешнюю антенну ФС II формируют субъединицы основного светособирающего комплекса ССК II, образующие триммеры Каждый мономер представлен тремя трансмембранными а-спиралями A, В и С, длина которых составляет 43 A, 51 A и 32 A, соответственно. «Функциональное ядро» или кор-комплекс ФС II (ПБК II) включает два антенных белка с М.м. 40-45 и 45-51 кДа (СР 43 и СР 47) и комплекс РЦ ФС II (D1-D2 {цит. b559 комплекс РЦ ФС II) Антенные белки связывают 13-15 молекул Xл. а и по две молекулы b-каротина. Если перенос энергии в пределах кластера осуществляется по экситонному механизму, то перенос энергии между кластерами на разных мономерах или между кластерами на разных тримерах во внешней антенне происходит по индуктивно-резонансному механизму вследствие больших расстояний между хромофорами.
Основная функция РЦ ФС II состоит в акцептировании электронов от кислородвыделяющего комплекса и дальнейшей передаче их в цепь переносчиков по направлению в ФС I. Первичным донором электрона в ФС II является фотоактивный пигмент Pб80, названный так по аналогии с 700 . Освещение вызывает выцветание полосы при 680 - 690 нм и некоторое увеличение поглощения при 820 нм, характерное для катион-радикала хлорофилла.
Большое внимание уделяют изучению природы первичного акцептора, рас-положенного между Pб80 и первичным пластохиноном. Оказалось, что дифферен-циальные спектры фотоиндуцированных изменений поглощения субхлоропластных фрагментов, обогащенных ФС II и измеренных в восстановительных условиях, соответствуют спектру фотовосстановления феофитина (Фф). Выцветание полос поглощения при 408{410, 422{428, 685 нм, а также полос при 518 и 545 нм, появле-ние широких полос при 450 и 650 нм, узкой полосы при 675 нм, характерны для появления Фф в реакции:
P*680Фф - P+680Фф.-
Восстановление P680обеспечивается вторичным донором тирозином, который получает электроны от водоотнимающего комплекса.
От феофетина электрон сбрасывается на хинон Qa, а с него на хинон Qb
Рисунок 3

А с хинонов – на пул пластохинонов
Рисунок 4

|
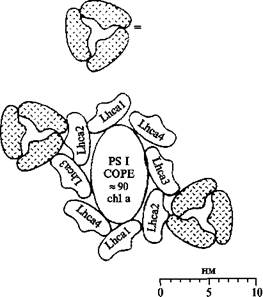
Комплекс ФСI
Рисунок 5
Кор-комплекс ФС I представляет собой гетеродимер, образованный большими белковыми субъединицами с М.м. 82,5 и 83,2 кДа, в состав которых входят как компоненты РЦ ФС I, так и антенные пигменты (около 100 молекул хл. a и 12 - 16 молекул b-каротина). Антенные пигменты кор-комплекса ФС I формируют по край-ней мере два кластера - коротковолновый (λ погл < 700 нм) и длинноволновый (так называемые «зацентровые» формы, λ погл > 700 нм), которые участвуют в переносе энергии на РЦ ФС I
Первичный донор P 700 электронов в ФС I был открыт в одном из первых экспериментов по применению дифференциальной оптической спектроскопии для исследования фотосинтетических мембран. Поглощение кванта света реакционным центром ФС I переводит P 700 в нижнее синглетное возбужденное состояние. Структурная и функциональная организация реакционного центра такова, что вероятность переноса электрона с возбужденного уровня на так называемый первичный фотохимический акцептор намного превышает вероятность излучательной или безизлучательной диссипации энергии этого возбужденного состояния. Локализовавшись на первичном акцепторе, электрон затем проходит по цепи переносчиков (вторичных акцепторов), окислительно-восстановительные потенциалы которых образуют нисходящую «лестницу» (соответствующую увеличению E, т.е. снижению восстановительной способности). Наиболее близким к Р700 непосредственным акцептором электронов от первичного донора служит, по-видимому, мономер хлорофилла a, полоса поглощения которого находится при 693{695 нм (Хл 695). Перенос электрона от 700 на Ao осуществляется быстрее чем за 10 пс и сопровождается выцветанием полосы Хл 695. После этого электрон за 20-50 пс переходит на акцептор A1, который, по-видимому, представляет собой филлохинон (витамин K 1 ). С каждым реакционным центром ФС I ассоциированы две молекулы филлохинона, одна из которых связана более прочно, чем другая.
Рисунок 6
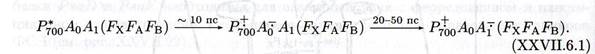
Восстановление P 700 происходит в цепи цит f → пластоцианин → Р+700 за время 200 мкс, а также частично непосредственно от цит f за 15 мкс
| <== предыдущая лекция | | | следующая лекция ==> |
| | | Динамическая подвижность белков |
Дата добавления: 2016-04-11; просмотров: 2583;