Ионные токи в модели Ходжкина-Хаксли. Воротные токи. Флуктуации напряжения и проводимости в модельных и биологических мембранах
Модель Х-Х – это математическая модель, описывающая электрофизиологические мембранные процессы возбудимых клеток. В частности, - гигантского аксона кальмара, с которым работали ученые. Математический аппарат модели строится на нескольких уравнениях. (более подробно – см. 12.1. тут все же вопрос про токи и проводимость)
1. Уравнение Нернста, описывающее равновесный потенциал в системе с полупроницаемой мембраной по каждому иону. φi=RT/(zF)* ln(Cin/Cout) (тут и далее под φ надо понимать разность потенциалов снаружи минус внутри. Дело в том, что правильнее говорить равновесная разность потенциалов, но поскольку привыкли говорить просто равновесный потенциал, то и вместо Δφ пишут φ, хотя по факу это неправильно). Оно выводится из условия равновесия, согласно которому электрохимические потенциалы в обеих частях системы, разделенной на эти части полупроницаемой мембраной, равны (µin= µ0+RTlnCin+zFφin и µout= µ0+RTlnCout+zFφout, µin= µout, писать с чертой!).
2. Уравнение Гольдмана-Ходжкина-Хаксли.Описывает потенциал на мембране и показывает, что он зависит от проницаемости мембраны по разным ионам (как оно выводится никогда не пишут в описании модели, вообще оно выводится с помощью электродиффузии, кому охота понтонуться – см. 3.1.) φ=RT/F*ln((PK[Kout]+ PNa[Naout]+ PCl[Clin])/( PK[Kin]+ PNa[Nain]+ PCl[Clout])).
3. 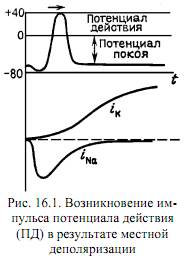 Уравнения для ионных токов, определяющих возникновение ПД и все его фазы. Ii=gi*(φ-φi), g – проводимость. (g=1/R). + если учесть, что в покое сумма всех токов равна нулю, то можно вывести следующее уравнение: φ=(gKφK+gNaφNa+gClφCl)/(gK+gNa+ gCl). Видно, что ток зависит от двух изменяющихся величин: потенциала на мембране и проводимости. Проводимость изменяется в зависимости от потенциала на мембране. Короче: Ii=f(φ, gi(φ)). φi – в данном случае константа. Известно, что gNa сначала быстро растет, а затем быстро экспоненциально убывает, а gK растет по S-образной кривой (см рис. 1 из 12.1). Вопрос о том, как gi управляется электрическим полем, вызванным потенцалом на мембране, - ключевой в биофизике мембранных процессов. В модели Х-Х предполагается, что проводимость регулируется некоторыми положительно заряженными частицами, которые перемещаются в мембране при изменении поля. Эти частицы соответсвующим образом закрывают или открывают ионным канал. Для К-каналов считается, что есть 4 активирующие частицы, а для Na-каналов 3 активирующие и 1 инактивирующая. Это возволило создать математический аппарат, с высокой точностью моделирующий нервный импульс. В функционально структуре канала удалось выделить элементы, отвечающие за механизмы селекции ионов – селективный фильтр, - активации – активационные ворота – и инактивации – инактивационные ворота. Эти управляющие частицы называются воротными частицами. Их движение можно обнаружить эксперементально по возникновению т.н. воротных токов. Они появляются в результате смещения частиц в мембране под влиянием наложенного на мембрану электрического потенциала. Удалось обнаружить воротные токи смещения, связанные с частицами, открывающими Na-канал. Предполагается, что эти токи вызваны изменением ориентации диполей, выстилающих внутреннюю полость канала. (см. рис. 2 из 12.1)
Уравнения для ионных токов, определяющих возникновение ПД и все его фазы. Ii=gi*(φ-φi), g – проводимость. (g=1/R). + если учесть, что в покое сумма всех токов равна нулю, то можно вывести следующее уравнение: φ=(gKφK+gNaφNa+gClφCl)/(gK+gNa+ gCl). Видно, что ток зависит от двух изменяющихся величин: потенциала на мембране и проводимости. Проводимость изменяется в зависимости от потенциала на мембране. Короче: Ii=f(φ, gi(φ)). φi – в данном случае константа. Известно, что gNa сначала быстро растет, а затем быстро экспоненциально убывает, а gK растет по S-образной кривой (см рис. 1 из 12.1). Вопрос о том, как gi управляется электрическим полем, вызванным потенцалом на мембране, - ключевой в биофизике мембранных процессов. В модели Х-Х предполагается, что проводимость регулируется некоторыми положительно заряженными частицами, которые перемещаются в мембране при изменении поля. Эти частицы соответсвующим образом закрывают или открывают ионным канал. Для К-каналов считается, что есть 4 активирующие частицы, а для Na-каналов 3 активирующие и 1 инактивирующая. Это возволило создать математический аппарат, с высокой точностью моделирующий нервный импульс. В функционально структуре канала удалось выделить элементы, отвечающие за механизмы селекции ионов – селективный фильтр, - активации – активационные ворота – и инактивации – инактивационные ворота. Эти управляющие частицы называются воротными частицами. Их движение можно обнаружить эксперементально по возникновению т.н. воротных токов. Они появляются в результате смещения частиц в мембране под влиянием наложенного на мембрану электрического потенциала. Удалось обнаружить воротные токи смещения, связанные с частицами, открывающими Na-канал. Предполагается, что эти токи вызваны изменением ориентации диполей, выстилающих внутреннюю полость канала. (см. рис. 2 из 12.1)

Полярные группы, определяющие диполыюе окружение иона в канале, влияют на энергию иона и на прохождение его через канал, Если иод влиянием деполяризующего электрического импульса изменяется их ориентация, то это вызовет смещение заряженных групп и изменение проводимости канала. Процесс переориентации диполей может носить кооперативный характер и быть достаточно резким. В этом случае энергия, необходимая для переориентации каждого элементарного диполя, должна зависеть не только от его собственной энергии, но и от доли диполей, уже изменивших свою ориентацию. Иными словами, по мере изменения ориентации части диполей энергия, необходимая для переориентации оставшихся диполей, уменьшается тем значительнее, чем больше число уже переориентированных диполей. Можно представить себе, что в исходном состоянии диполи мешают друг другу изменить ориентацию под действием поля, а переориентированные диполи уже "не путаются иод ногами" у оставшихся. В результате такого рода кооперативного эффекта проводимость канала очень быстро "лавинообразно" нарастает под действием приложенного электрического импульса. Заметим, что и конформацнонные перестройки в канале, сопровождающиеся поворотом диполей, могут также приводить к скачкообразным изменениям проводимости в одиночном канале. Фактически во всех предложенных моделях речь идет о своего рода фазовых переходах в мембранах, лежащих в основе скачкообразных переходов канала между двумя состояниями.
Короче, суть в том, что на самом деле, в основе модели лежат как раз дико сложные диффуры, которых не было у нас в задачах, и которых нет в лекциях, но которые есть в описаниях модели на стороне. И там как раз страшные диффуры про все эти изменения проводимостей и токов, но они реально страшные... Так что, извините, но не вижу смысла них разбираться, то есть тратить ден на то, чего все равно даже самый заядлый извращенец не спросит.
Тем не менее, опишу, какие токи учитываются в модели, кроме обычных калиевых и натриевых. Не знаю, насколько мы это должны знать... Нам про эти токи очень много рассказывали на нейрофизиологии, но на биофизике – едва ли... Кто хочет понтонуться.
Калиевые токи.
1.1.А-ток. У нейронов беспозвоночных деполяризация мембраны активирует новый тип К-канала, который, в отличие от К-канала в аксоне инактивируется со временем при продолжающейся деполяризации мембраны. Включение такого быстро инактивирующегося К-тока в модель генерации ПД позволяет модифицировать скорость (или частоту ритмического возбуждения) генерации ПД. Такой К -ток называют А-ток. Он быстро инактивируется и не активируется до тех пор, пока не произойдет кратковременная гиперполяризация мембраны. Активируется А-ток во время спада "обычного" калиевого тока, когда потенциал мембраны составляет ~ -75 мВ. А-ток предотвращает деполяризацию и т.о. снижает частоту импульсного разряда. Приложение электрического тока к нервным клеткам симпатических ганглиев лягушки-быка приводит к генерации сложного вида ПД из-за активации большого количества разных каналов. Присутствует по крайней мере 4 различных К+-тока: помимо А-тока могут быть выделены также два тока, которые активируются при поступлении в клетку Са++ во время ПД (IC и IAPH) и один, который активируется при деполяризации (IM) мембраны.
1.2.С-ток. Реполяризация аксолеммы в процессе ПД обеспечивается как инактивацией Nа-тока, так и активацией К-тока. Однако, для многих типов нервных клеток было показано, что удаление внеклеточного Са++ или блокирование Са++-каналов двухвалентными катионами(например Cd++, для которых мембрана непроницаема), приводит к удлинению фазы реполяризации и снижению уровня гиперполяризации мембраны после прохождения ПД. Эти результаты указывают на то, что Са2+ может транспортироваться в клетку в течение ПД и в связи с этим происходит активация К+-тока (IC), обеспечивающая реполяризацию мембраны.
1.3.АНР-ток. Для ряда нейронов выявлен другой тип Са2+-активируемого К+-тока. Например, внутриклеточная деполяризация пирамидальных нейронов неокортекса человека импульсом тока приводит к генерации серии ПД, сопровождающейся следовой гиперполяризацией клеточной мембраны. Такая следовая гиперполяризация - главным образом результат активации медленного К+-тока, IAHP (after hyperpolarisation), который генерируется за счет входа Са2+ при проведении серии ПД. При генерации ПД Са2+ поступает в клетку через Са2+-каналы, активирующиеся при высоких напряжениях стимулирующего клетку импульса. Увеличение количества внутриклеточного Са2+ активирует IAHP, который затем гиперполяризует мембрану клетки и приводит к уменьшению частоты генерации ПД (процесс известен как «частотная адаптация». Активация различных рецепторов, включая ацетилхолиновые (мускариновые) рецепторы, локализованных на мембране пирамидальных клеток коры, уменьшает IAHP и, соответственно, повышает частоту генерации ПД. Данное увеличение возбудимости пирамидальных клеток коры является важным фактором, обеспечивающим поддержание активности коры мозга во время бодрствования; и, наоборот, клетки коры становятся менее возбудимыми в состоянии сна без сновидений.
1.4.М-ток. Генерация разнообразных пачек ПД в нейроне осуществляется за счет взаимодействия различных ионных токов, которые могут меняться при действии нейромедиаторов. Одним из примеров такого типа нейромодуляции был обнаружен в клетках симпатических ганглиев лягушки-быка Д. Брауном и П. Адамсом. Эти исследователи выявили механизмы, с помощью которых высвобождение медиатора из окончаний предганглионарных нейронов спинного мозга (ацетилхолин) модулирует возбудимость ганглионарных клеток за счет активации мускариновых рецепторов. Амплитуда медленного возбуждения (деполяризации) обусловлена активацией мускариновых рецепторов при высвобождении ацетилхолина и зависит от МП клетки. Браун и Адамс продемонстрировали, что потенциалозависимость медленного холинэргического возбуждения обусловлена инактивацией ацетилхолином потенциалозависимого К+-тока (М-ток), который блокирует мускариновые рецепторы. Таким образом, М-ток - это К+-ток, активирующийся при деполяризации клетки за время порядка десятка миллисекунд. Например, если мы с помощью метода фиксации потенциала изменим МП клетки с -65 мВ на -45 мВ, то М-ток постепенно включится. Стимуляция предганглионарного нерва при действии ацетилхолина приводит к значительному подавлению этого медленно активирующегося К+-тока. Активация М-тока при деполяризации мембраны приводит к снижению клеточной возбудимости настолько, что прикладываемый к нерву деполяризующий импульс тока способен генерировать не пачку, а лишь один ПД. Однако, при блокировании М-тока в момент выхода из нейрона ацетилхолина, аналогичный по величине деполяризующий импульс приводит к генерации пачки ПД.
Кальциевые токи.
При исследовании состояния нейронов мозга спящих животных было обнаружено, что многие клетки (даже во время фазы медленного сна) не находятся в состоянии покоя, а генерируют серии ПД, а не отдельные ПД (типично для бодрствующих животных). Действительно, внутриклеточная регистрация МП во время фазы медленного сна для таламических клеток, которые осуществляют передачу сигналов от сетчатки к зрительной коре, выявила наличие медленных ПД, обеспечивающих серии импульсов в спящем мозге. Переход к бодрствованию или быстрому сну был связан с деполяризацией мембраны и более регулярной генерацией ПД в отличие от спайков или серии ПД. Дальнейшие исследования доказали наличие изменений ионных токов нейронов при переходе животного от состояния сна к бодрствованию. С помощью микроэлектродных исследований, выполненных на таламических клетках, были выявлены два основных способа генерации ПД. В случае, когда мембрана гиперполяризована, внутриклеточное введение в клетку деполяризующего импульса тока (до значений <-60 мВ) приводило к активации низкопорогового Са -зависимого ПД, что стимулировало серию (пачку) из 3-5 ПД. Однако, если клеточный МП составлял -63 мВ, внутриклеточное приложение такого же импульса тока приводит только к пассивному ответу клетки, который определяется емкостью и сопротивлением ее мембраны. Если плазматическая мембрана клетки затем деполяризуется до -53 мВ, то аналогичный импульс приводит к генерации обычной серии ПД, при это частота, в отличии от пирамидальных клеток коры, со временем не уменьшается. Таким образом, даже на уровне отдельных нейронов проявляются изменения активности в отношении цикла сна и бодрствования.
Наличие у таламических клеток механизма генерирующего пачки ПД во время медленного сна обусловлено свойствами особого типа Са2+-тока, известного как низкопороговый, или быстрый, Са2+-ток, обозначаемый также как Т-ток. Медленный ПД возникает при входе Са2+ в клетку. Как и Nа+-ток, участвующий в генерации ПД, Т-ток способен к инактивации, однако он инактивируется уже при деполяризации до ~-60 мВ, и длительная деполяризация клетки приводит к полной его инактивации и исчезновению медленных спайков. Эта особенность Т-тока обеспечивает способность таламических нейронов переходить от состояния генерации вспышек ПД во время сна к нормальной генерации ПД при состоянии бодрствования.
Высокопороговый Са2+-ток (L-ток), активируемый только при значениях более МП ~ - 30мВ и медленно инактивируемый при деполяризации, участвует в активации Са2+-активируемого К+-тока.
| <== предыдущая лекция | | | следующая лекция ==> |
| Многие молекулы биологических веществ являются природными или естественными флуорофорами | | | Технологиялық сүлбесі |
Дата добавления: 2016-04-11; просмотров: 3101;