Уздзеянне iанiзуючага выпраменьвання на лясныя бiягеацэнозы
12.7.1. Асаблiвасцi лясных бiягеацэнозаў як аб'ектаў радыяцыйнага забруджвання
Цiкавасць да ляснога бiягеацэнозу як аб'екта радыяцыйнага забруджвання тлумачыцца шырокай распаўсюджанасцю лясоў на зямным шары i той сапраўды планетарнай роляй, якую яны выконваюць у падтрыманнi кiслароднага балансу ў атмасферы i воднага балансу сушы, ахове глебы ад эрозii, у велiзарным народнагаспадарчым значэннi лесу як пастаўшчыка драўнiны i iншых вiдаў лясной прадукцыi.
Лес уяўляе сабой своеасаблiвы прыродны комплекс, у якiм першапачатковае размеркаванне i далейшая мiграцыя радыенуклiдаў маюць сваю спецыфiку ў параўнаннi з iншымi тыпамi ландшафтаў. Ён мае здольнасць трывала ўтрымлiваць радыенуклiды, прадухiляць iх вынас за межы забруджанай тэрыторыi. Наяўнасць на шляху гарызантальных паветраных патокаў буйных масiваў лесу i нават асобных дрэў прыводзiць да затрымання на ствалах, iглiцы, лiсцi i iншых паверхнях радыеактыўнай аэразолi, што пераносіцца гэтымi патокамi. У вынiку яруснага размяшчэння крон i вялiкай паверхнi асiмiлюючых органаў дрэвавай раслiннасцi значная частка выпаўшых радыенуклiдаў затрымлiваецца. Больш высокая канцэнтрацыя радыенуклiдаў назiраецца ў наветраных частках крон дрэў. Турбулентны рух паветраных мас садзейнiчае раўнамернаму вертыкальнаму размеркаванню радыенуклiдаў у полагу лесу. Радыенуклiды, якiя асядаюць з атмасферы на кроны высокапаўнотных хваёвых лясоў, могуць затрымлiвацца амаль поўнасцю.
Другiм раслiнным фiльтрам для асядаючых радыенуклiдаў з'яўляецца травяны ярус, затрымлiваючая здольнасць якога залежыць ад фiтамасы, памераў, ажурнасцi экспануючай паверхнi i г. д.
У залежнасцi ад хуткасці ветру iнтэнсiўнасць максiмальнага асядання радыенуклiдаў у лясах можа быць пры розных метэаралагiчных умовах у 6–12 разоў вышэйшай, чым у травяністай раслiннасцi. Змяншэнне ўтрымання радыенуклiдаў у аблоках пры руху iх над лесам адбываецца хутчэй, чым над прасторай з травянiстай раслiннасцю. Гэта сведчыць аб тым, што лес з'яўляецца дастаткова эфектыўным фiльтрам для радыенуклiдаў. Аднак павышаная сорбцыя радыенуклiдаў ляснымi бiягеацэнозамi служыць крынiцай узмоцненага iх апраменьвання, а пры высокiх паглынутых дозах – прычынай iх радыяцыйнага паражэння.
Iнфармацыя аб забруджваннi лясных бiягеацэнозаў з'яўляецца неабходнай i дастатковай умовай для разлiку паглынутых доз, ролi лесу як санiтарна-гiгiенiчнага фактару, якi ахоўвае тэрыторыi i iх жыхароў ад другаснага радыеактыўнага забруджвання.
Значную цiкавасць маюць звесткi аб здольнасцi лясных арганiз-маў, у першую чаргу дрэвавых i травянiстых раслiн, да назапашвання доўгажывучых прадуктаў дзялення, якiя паступаюць з глебы. Колькас-ныя даныя аб назапашваннi, размеркаваннi i мiграцыi радыенуклiдаў у лясных насаджэннях служаць навуковай асновай для распрацоўкi практычных рэкамендацый, рацыянальных метадаў вядзення лясной гаспадаркi на тэрыторыях з павышаным узроўнем радыеактыўнага забруджвання.
Колькаснай характарыстыкай першаснага ўздзеяння радыенуклiдаў з'яўляецца каэфiцыент затрымання (Кпз), якi вызначаецца як адносiны колькасцi радыенуклiда, першапачаткова затрыманага раслiнным покрывам (А1, Кi/км2), да колькасцi радыенуклiда першапачаткова затрыманага адзiнкай паверхнi (А0, Кi/км2). Велiчыня гэтага каэфiцыента залежыць ад тыпу лесу i ўзросту дрэвастою, метэаралагiчных умоў i фiзiка-хiмiчнай формы выпаўшых радыенуклiдаў i вагаецца ў шырокiх межах.
Да фактараў, якiя аказваюць моцны ўплыў на бiягеацэнозы, адносяцца i iанiзуючыя выпраменьваннi. Яны парушаюць iснуючыя сувязi i могуць прывесцi да лятальнага зыходу або адмiрання спачатку найбольш, а затым i найменш, радыеадчувальных вiдаў.
Узаемасувязi памiж кампанентамi ляснога бiягеацэнозу ўключаюць трафiчныя адносiны розных груп арганiзмаў, прамыя i зваротныя сувязi дрэвавых раслiн з мiкраклiматам пад iх полагам, розныя вiды сiмбiёзу i г.д. У нармальных умовах гэтыя сувязi збалансаваны так, што лясны бiягеацэноз прадстаўляе сабой устойлiвую самарэгулявальную сiстэму, якая функцыянуе без неабарачальных парушэнняў на працягу многiх дзесяцiгоддзяў.
Каэфiцыент затрымання радыенуклiдаў ярусам вечназялёных дрэвавых раслiн прымаюць роўным ступенi самкнутасцi крон, лiстападных – у тры разы меншым. Пасля выпадання радыенуклiдаў пачынаецца вертыкальная i гарызантальная iх мiграцыя пад уздзеяннем ветру, ападкаў i з ападам, у вынiку чаго яны перамяшчаюцца з верхнiх частак крон у нiжнiя, а затым – пад полаг лесу (табл. 12.4).
Таблiца 12.4
Размеркаванне паглынутых доз g-выпраменьвання (Гр/сут) па вертыкалi хваёвага лесу пры радыеактыўным выпаданнi з атмасферы сумесi радыенуклiдаў ( 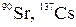 ) 109 Бк/м2 у асеннi перыяд (паводле Крывалуцкага Д. А. i iнш.)
) 109 Бк/м2 у асеннi перыяд (паводле Крывалуцкага Д. А. i iнш.)
| Прайшло з моманту выпадання, месяцаў | Паверхня глебы | Паверхня подсцiлу, 2 см | Нiжняя частка крон, 3 м | Сярэдняя частка крон, 6–7 м | Верхняя частка крон, 9–10 м |
| 0,04 | 0,16 | 1,3 | 1,8 | 1,0 | |
| 1,0 | 5,6 | 2,0 | 1,0 | 0,7 | |
| 1,4 | 7,0 | 0,8 | 0,9 | 0,5 | |
| 1,4 | 6,2 | 0,5 | 0,4 | 0,2 | |
| 3,7 | 2,4 | 0,15 | 0,05 | 0,003 | |
| 5,6 | 1,2 | 0,08 | 0,03 | 0,015 |
Шматгадовыя даследаваннi Крывалуцкага Д.А. i iнш. па кругавароце 90Sr у спелых бярозавых i сасновых лясах паказалi, што працягласць этапу, за якi каля 90% радыенуклiда перамяшчаецца пад полаг бярозавага лесу, складае каля года, у хваёвых – 3–5 гадоў. Пасля завяршэння гэтага этапа асноўнай крынiцай паступлення радыенуклiда ў надземную частку дрэвастою з'яўляецца глеба. Дынамiка ўтрымання 90Sr у бiямасе сасны i бярозы паказана на рыс. 12.3.
| |
| |
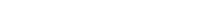

 |
Рыс. 12.3. Дынамiка ўтрымання 90Sr у бiямасе сасны (а) i бярозы (б) у 30-гадовай саснова-бярозавай пасадцы пры шчыльнасцi забруджвання 1 МБк/м2:
а: 1 – флаэма; 2 – коркавы слой кары; 3 – галлё тонкае; 4 – iглiца; 5 – драўнiна; б: 1 – лiсце; 2 – флаэма; 3 – бяроста; 4 – галлё; 5 – драўнiна
Праз чатыры гады пасля ўнясення радыенуклiда ў глебу ў кампанентах дрэў бярозы ўтрыманне 90Sr стабiлiзавалася i на працягу некалькiх гадоў заставалася практычна нязменным. У тонкiм галлi i iглiцы сасны на працягу шасцi гадоў утрыманне 90Sr рэзка змяншалася, а пасля перамяшчэння радыенуклiда на паверхню глебы зноў павялiчвалася.
Пасля перамяшчэння асноўнай часткi радыенуклiдаў пад полаг крон значныя дозы апраменьвання атрымлiваюць насельнiкi нiжнiх ярусаў лесу. У першыя два месяцы пасля паступлення (рыс. 12.4) асноўная колькасць 90Sr знаходзiлася ў верхнiм пласце глебы. Праз 13 гадоў значная частка радыенуклiдаў перамясцiлася па профiлi на глыбiню да 5–8 см. 137Cs за шэсць гадоў мiгрыраваў толькi на глыбiню да 2 см.
| |
|

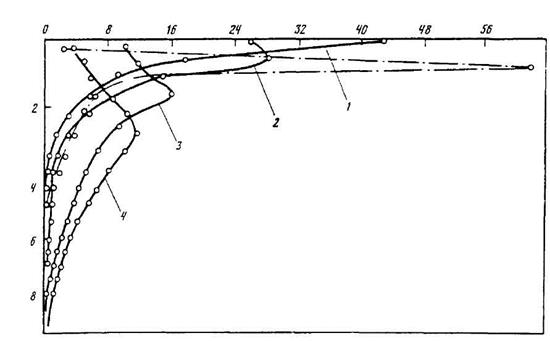
Рыс. 12.4. Размеркаванне па профiлi шэрай лясной глебы 137Cs (пункцiрная лiнiя) праз 6 гадоў i 90Sr (суцэльная лiнiя) праз 2 месяцы (1), 2 гады (2), 8 гадоў (3) i 13 гадоў (4) пасля ўнясення радыенуклiдаў на паверхню ляснога подсцiлу
Дрэвы з часткова пашкоджаным камбiем маюць нiзкую ветраўстойлiвасць у вынiку парушэння структуры драўнiны i яе адмiрання ў месцах гiбелi камбiю. Большая частка такiх дрэў на працягу двух гадоў ламаецца. На рыс. 12.5 добра бачна дэструктыўнасць драўнiны на рознай вышынi ствала. Мёртвая частка размешчана з боку пашкоджання камбiю. Пры пэўных умовах апраменьвання крытычнай тканкай дрэў могуць быць не пункты росту, а латэральная мерыстэма камбiю – з-за неаднолькавага размеркавання паглынутых доз па вертыкалi насаджэння.
 |
Рыс. 12.5. Зрэзы ствала бярозы (паглынутая доза ў ствале 1000 - 1500 Гр) узятыя на вышынi:

|
|
|
|
|
|
|
|
12.7.2. Радыеадчувальнасць асноўных кампанентаў лясных бiягеацэнозаў
Радыебiялагiчным аналiзам кампанентаў лесу выяўлена надзвычай вялiкая амплiтуда (да 500 разоў) радыеадчувальнасцi раслiн i жывёл, якiя насяляюць прыродныя бiягеацэнозы розных прыродных зон зямнога шара. Прычыны рознай радыеадчувальнасцi асобных вiдаў найбольш поўна тлумачыць тэорыя эколага-геаграфiчнага паходжання вiдаў (Кудзiнаў М. А.). Гэтая канцэпцыя грунтуецца на тым, што ў раслiн адсутнічаюць спецыяльныя ахоўныя прыстасаванасцi да такога экалагiчнага фактара навакольнага асяроддзя, як пранiкальная радыяцыя, а комплекс прыстасаванасцi дзейнiчае як засцярога.
Даследаваннямi прамянёвага пашкоджання лясоў выяўлена складанае ўзаемадзеянне радыяцыйнага фактару з iншымi ўмовамi навакольнага асяроддзя (цеплавым, водным, светлавым i iнш.). У эксперыментах з апраменьваннем лясных бiягеацэнозаў атрыманы звесткi аб марфалагiчных, фiзiялагiчных, цытагенетычных i iншых змяненнях. У апошнiя гады супрацоўнiкамi Iнстытута лесу i Iнстытута эксперыментальнай батанiкi НАНБ, ЦБС НАНБ i iншых у лясах рэспублiкi з рознай ступенню ўздзеяння iанiзуючага выпраменьвання праводзяцца даследаваннi ўплыву выпраменьвання на асобныя кампаненты лясных бiягеацэнозаў.
Найбольш радыеадчувальнымi з'яўляюцца кампаненты лясных бiягеацэнозаў, якiя адрознiваюцца павышанай сорбцыяй радыенуклiдаў (табл. 12.5). Гэта пацвярджае неабходнасць больш дэталёвага вывучэння ўплыву малых доз iанiзуючых выпраменьванняў на лясныя бiягеацэнозы i асобныя iх кампаненты.
Таблiца 12.5
Класiфiкацыя прыродных бiягеацэнозаў па радыеадчувальнасцi
(па Аляксахiну Р. М.)
| Тып бiягеацэноза | Дозы, якiя вядуць да прамянёвага пашкоджання, Гр | ||
| слабага | сярэдняга | моцнага | |
| Хваёвы | 1–10 | 10–20 | >20 |
| Мяшаны | 10–100 | 50–350 | 100–600 |
| Трапiчны дажджавы | 40–100 | 100–400 | >400 |
| Кустовы | 10–50 | 50–200 | >200 |
| Лугавы | 80–100 | 100–1000 | >1000 |
| Мохава-лiшайнiкавы | 100–500 | 500–5000 | >5000 |
| Лiшайнiкавы | 600–1000 | 1000–2000 | >2000 |
Розная радыеадчувальнасць кампанентаў лясных бiягеацэнозаў у зонах радыеактыўнага забруджвання ў канчатковым вынiку прыводзiць да збяднення вiдавой разнастайнасцi, змянення хiмiчнага саставу i фiзiка-хiмiчных уласцiвасцей подсцiлу i глебы, рэпрадукцыйных, роставых i iншых працэсаў. Самы вялiкi ўплыў на лясную раслiннасць аказваюць 137Cs i 90Sr.
12.7.3. Фактары, якія ўплываюць на радыеадчувальнасць і радыеўстойлівасць раслін
Уплыў іанізуючага выпраменьвання на асобныя кампаненты лясных біягеацэнозаў залежыць ад шматлікіх фактараў: тыпу выпраменьвання, тыпу і працягласці апраменьвання, дысперснасці радыеактыўных часціц, генетычных і фізіялагічных асаблівасцей, экалагічных фактараў, пары года, характарыстыкі біягеацэнозаў і інш.
Тыпы выпраменьвання. Пры знешнім апраменьванні раслін і жывёл самы вялікі ўплыў аказвае γ-выпраменьванне, якое адрозніваецца высокай пранікальнай здольнасцю. Значны радыебіялагічны эфект α- і β-выпраменьвання назіраецца тады, калі яны пападаюць унутр раслін праз вусцейкі або каранёвыя сістэмы з глебавым растворам і з ежай у арганізмы жывёл. α-часціцы пашкоджваюць слізістыя абалонкі ўнутраных органаў. Нязначны ўплыў α- і β-часціцы аказваюць на расліны і жывёл у тым выпадку, калі затрымліваюцца пакроўнымі тканкамі і скурным покрывам.
Пры хранічным апраменьванні раслін і жывёл аднолькавымі дозамі радыеадчувальнасць і радыеўстойлівасць залежаць ад генетычных фактараў і стану арганізма. Даследаваннямі ўстаноўлена, што па меры ўскладнення арганізма радыеадчувальнасць яго ўзрастае. Чым большы аб’ём у клетачных ядрах займаюць храмасомы і чым большы аб’ём ядраў, тым у большай ступені яны падвяргаюцца храмасомным аберацыям.
Пры фракцыянаваным апраменьванні арганізм паспявае часткова або поўнасцю аднавіцца пасля першага апраменьвання, а яго радыеадчувальнасць можа павялічвацца або змяншацца, што залежыць ад дозы.
Кропкавае апраменьванне высокімі дозамі выклікае гібель толькі асобных органаў або іх частак. Пры гэтым жыццядзейнасць усяго арганізма захоўваецца. Малыя дозы могуць выклікаць паляпшэнне паказчыкаў росту асобных органаў і тканак.
Дысперснасць радыеактыўных часціц. Больш дробныя часціцы радыенуклідаў пры аднолькавай іх масе аказваюць значны ўплыў на радыеадчувальнасць і радыеўстойлівасць арганізмаў. Гэта звязана з павелічэннем паверхні, на якую ўздзейнічаюць радыеактыў-ныя часціцы. Павелічэнне працягласці ўздзеяння радыенуклідаў на ар-ганізмы ва ўсіх выпадках прыводзіць да зніжэння радыеўстойлівасці.
Фізіялагічныя фактары. На радыеадчувальнасць арганізмаў значны ўплыў аказвае перыяд развіцця, індывідуальныя асаблівасці, перыяд вегетацыі, узрост і іншыя фактары. Вядома, што ў розныя перыяды антагенезу арганізмы як раслін, так і жывёл адрозніваюцца не толькі марфалагічнай будовай, але і спецыфікай фізіка-хімічных працэсаў і радыеадчувальнасцю. Максімальная яна ў насення ў перыяд яго прарастання і пераходу раслін ад вегетатыўнага да генератыўнага стану, а самая вялікая радыерэзістэнтнасць – у перыяд паспявання і ў стане пакою.
Экалагічныя фактары. Расліны і жывёлы прызвычайваюцца да тых экалагічных умоў, у якіх яны працяглы час знаходзяцца. Рэзкае змяненне выклікае тыя ці іншыя рэакцыі арганізмаў і нават поўную іх гібель. Павышэнне ўтрымання радыенуклідаў у асноўным адмоўна ўплывае на стан арганізмаў. Найбольш нізкая радыеадчувальнасць дрэвавых відаў назіраецца ў асенне-зімовы перыяд, высокая – у час распускання пупышак.
12.7.4. Аэральнае i каранёвае паступленне радыенуклiдаў
У раслiны
Аэральнае паступленне радыенуклiдаў у раслiны адбываецца пры асяданнi iх на надземныя часткi кампанентаў біягеацэнозаў аэразольных i газападобных выкiдаў. Крынiцамi забруджвання могуць быць лакальныя i глабальныя выпадзеннi пры выпрабаваннi ядзерных установак, нармальныя або аварыйныя выкiды радыенуклiдаў пры рабоце прадпрыемстваў на ядзерным палiве.
Рэгулярныя i аварыйныя выкiды АЭС складаюцца з iнертных газаў, лятучых i нелятучых радыенуклiдаў i iнш. Значная частка прадуктаў дзялення i нуклiдаў з наведзенай актыўнасцю ўтварае аэразоль, якая i выводзiцца ў навакольнае асяроддзе з паветранымi выкiдамi. Пры падземных i падводных ядзерных выбухах могуць узнiкаць сiтуацыi, пры якiх ствараюцца ўмовы для пападання радыенуклiдаў у атмасферу. У асноўным гэта iнертныя газы, а таксама радыеактыўная аэразоль з газападобных папярэднiкаў, якiя ўтрымлiваюць 89Sr, 90Sr, 137Cs. Важным кампанентам радыеактыўных выкiдаў у атмасферу могуць быць радыенуклiды, якiя дрэнна сарбiруюцца фiльтравальнымi прыстасаваннямi ў месцах iх выкiду. Для некаторых з гэтых радыенуклiдаў пазакаранёвае паступленне з'яўляецца асноўным шляхам уцягнення ў ланцугi мiграцыi.
Радыенуклiды, што паступаюць у атмасферу, утвараюць аэразоль i пад уплывам гравiтацыйных сiл, а таксама пад уздзеяннем метэаралагiчных фактараў (дождж, снег, туман) выпадаюць на паверхню зямлi. Асяданне радыенуклiдаў на раслiннае покрыва адбываецца i ў днi без ападкаў. Часам доля сухiх выпадзенняў вельмi вялiкая. Гэтаму садзейнiчае турбулентны рух паветраных мас у атмасферы.
Выпаданне радыеактыўнай аэразолi на паверхню раслiн прыводзiць да назапашвання ў iх надземнай фiтамасе ўсёй сукупнасцi асеўшых радыенуклiдаў, у той час як пры каранёвым шляху паступлення iх у раслiны глебавы паглынальны комплекс выступае ў ролi магутнага сарбцыйнага фактару, а каранёвыя сiстэмы раслiн з'яўляюцца селектыўным бар'ерам, якi выключае паступленне ў надземную масу бiялагiчна-iнертных элементаў.
Радыенуклiды могуць паступаць у раслiны ў вынiку падняцця ветрам або кроплямi дажджу з наглебавага покрыва як самiх радыеактыўных часцiнак, так i забруджаных крупiнак глебы. Гэта з'ява называеццадругасным забруджваннем раслiн.
Радыенуклiды, якiя аселi з паветра i затрымалiся на паверхнi лiсцяў (iглiцы), галля, ствалоў дрэў, кустоў i травяна-імховым покрыве, могуць знаходзiцца як у свабодным, так i ў сарбiраваным стане. На сорбцыю затрыманых раслiнамi радыенуклiдаў аказвае ўплыў тэмпература i вiльготнасць паветра, асаблiвасцi будовы паверхнi лiсцяў, наяўнасць або адсутнасць кутыкулы, салявы састаў i рН атмасферных ападкаў, з якiмi на раслiны асядаюць радыенуклiды. Перамяшчэнне затрыманых раслiнамi радыенуклiдаў у органы, куды яны непасрэдна не паступалi, вызначаецца як фiзiка-хiмiчнымi ўласцiвасцямi радыенуклiдаў, так i бiялагiчнымi асаблiвасцямi раслiн.
Перамяшчэнне i замацаванне радыенуклiдаў у верхнiх гарызонтах глебы забяспечвае iснаванне ў прыродзе дзеючай крынiцы для каранёвага назапашвання iх раслiнамi. Моцная сорбцыя радыенуклiдаў цвёрдай фазай глебы абмяжоўвае iх засваенне праз каранёвыя сiстэмы. Паглынанне радыеактыўных рэчываў раслiнамi з глебы, у першую чаргу, залежыць ад яе ўласцiвасцей i ад глебавай хiмii радыенуклiдаў.
Для ацэнкi паступлення радыенуклiдаў з глебы ў раслiны выкарыстоўваюць розныя паказчыкi. Адным з iх з'яўляецца каэфiцыент назапашвання радыенуклідаў Кн (або каэфiцыент канцэнтрацыi Кк), якi паказвае адносiны ўтрымання радыенуклiда ў адзiнцы масы раслiн i глебы адпаведна. Блiзкiм да паняцця Кн з'яўляецца глебавы каэфiцыент прапарцыянальнасцi Кп, якi суадносiць канцэнтрацыю радыенуклiдаў у раслiнах і паглынутае забруджванне глебы.
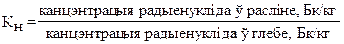
Замест паняцця каэфiцыент назапашвання выкарыстоўваюць каэфiцыент бiялагiчнага паглынання Кбп з дапамогай якога параўноўваюць канцэнтрацыю радыенуклiда ў попеле раслiн i глебе.
Асаблiвасцi мiнеральнага засваення, розная працягласць вегетацыйнага перыяду, характар размяшчэння каранёвых сiстэм у глебе, адрозненне ў прадукцыйнасцi i iншыя бiялагiчныя асаблiвасцi раслiн уплываюць на назапашванне радыенуклiдаў рознымi вiдамi. Адрозненнi ў акумуляцыi радыенуклiдаў асобнымі відамі пры каранёвым шляху іх пераходу могуць дасягаць 10–30 разоў.
12.7.5. Уплыў iанiзуючага выпраменьвання на фiзiялагiчныя працэсы i рост дрэў
Да гэтага часу ўжо назапашаны значны эксперыментальны матэрыял, што сведчыць аб уплыве iанiзуючых выпраменьванняў на розныя кампаненты, i ў тым лiку на дрэвавы ярус, якi складае аснову ляснога бiягеацэнозу. Парушэннi, якiя адбываюцца ў гэтым ярусе i iншых кампанентах у вынiку апраменьвання, выклiкаюць адмоўныя i дадатныя змяненнi ў працяканнi фiзiялагiчных працэсаў, наступленнi феналагiчных фаз, у генератыўных органах, iглiцы i лiсцi, роставых пупышках, росце i развiццi, адмiраннi i ўсыханнi дрэў i дрэвастояў.
Наступленне феналагiчных фаз. Характэрнай рэакцыяй на апраменьванне з'яўляюцца зрухi ў наступленнi асноўных фенафаз у развiццi дрэў. Назiраецца запавольванне ў распусканнi лiсця i больш раннi лiстапад. Такiя эфекты апраменьвання былi зарэгiстраваны Карабань Р. Т., Мiшанковым Б. С. i iнш., Карабань Р. Т., Спiрыным Д. А. i iншымі ў розных эксперыментах па ўплыве iанiзуючага выпраменьвання на лес. У лiставых парод (бяроза, асiна) у першую пасля апраменьвання вегетацыю пры паглынутых дозах да 1000 Гр асенняе пажаўценне лiсця наступала на 20–30, а ападанне – на 13–14 дзён раней. У наступныя пасля апраменьвання гады феналагiчнае развiццё апрамененых дрэў працякала без iстотных змяненняў. У дрэў сасны пры паглынутых дозах звыш за 10 Гр у год апраменьвання назiралася затрыманне пачатку росту верхавiнкавых i бакавых парасткаў на 8–10 дзён i спазненне цвiцення – на 2–3 днi. Пры дозах вышэй 5 Гр назiралася больш ранняе ападанне iглiцы старэйшага ўзросту. Пры ўзроўнi забруджвання глебы 5–25 Кi/км2 назiралася паскарэнне цвiцення i паспявання насення (Цiхамiраў Ф. А.).
Змяненне генератыўных органаў. Пашкоджанне iглiцы i лiсця адзiн з найбольш небяспечных прамянёвых эфектаў у лясных бiягеацэнозах, таму што з гiбеллю лiставага апарату звязана пашкоджанасць i гiбель дрэў. Найбольш глыбокi аналiз марфагенезу ў сасны i елкi пад уплывам iанiзуючага выпраменьвання, выклiканага катастрофай на ЧАЭС, прыведзены Козубавым Г. М., Градзiнскiм Д. М. i iнш. Да iх адносяцца змяненне формы органаў, парушэнне геатрапiзму, парадку галiнавання i iнш. Найбольш шырокi спектр марфалагiчных анамалiй назiраецца пры паглынутых дозах 3–4 Гр. Парушэнне марфагенезу вегетатыўных органаў у сасны часцей за ўсё абумоўлена пашкоджаннем мерыстэмных тканак. Адмiранне верхавiнкавых мерыстэм выклiкае закладку шматлiкiх бакавых пупышак i парасткаў з хаатычнай арыентацыяй, што прыводзiць да шматвяршыннасцi дрэў. Зарэгiстраваны анатамiчныя i гiсталагiчныя адхiленнi.
У год катастрофы на ЧАЭС Козубаў Г. М. i А. I. Таскаеў А. I. пры сярэдняй пашкоджанасцi з паглынутай дозай 3–4 Гр у апiкальнай частцы верхавiнкавых парасткаў назiралi пакарочаную iглiцу ў выглядзе густых шчотак. Сустракалася iглiца павялiчаных памераў у 1,5–2,0 разы. У наступны год «гiганцкая» iглiца сустракалася практычна на ўсiх дрэвах з часткова пашкоджанымi кронамi. Клеткi мезафiлу гэтай iглiцы па памерах былi блiзкiмi да нормы. Большасць хларапластаў мела асобныя вырасты. Вострае апраменьванне з паглынутай дозай 80–100 Гр i больш прыводзiла да поўнага адмiрання надземных органаў сасны на працягу кароткага тэрмiну. Восенню 1986 i вясной 1987 гг. на некаторых знешне загiнуўшых соснах утваралiся i хутка ўсыхалi адзiнкавыя зялёныя парасткi.
Характэрнай асаблiвасцю пашкоджання сасны пры апраменьваннi дозамi 15–40 Гр з'яўлялася пажаўценне пучкоў iглiцы каля асновы верхавiнкавых i бакавых роставых пупышак на аднагадовых парастках двух-пяцi верхнiх кальчакоў дрэў i ападанне iх у пачатку другога года вегетацыi (Карабань Р. Т.). Пры дозе апраменьвання 40–60 Гр у верхняй частцы кроны сосен назiралася пажаўценне двухгадовай iглiцы на 1/2–3/4 даўжынi парасткаў. На астатняй частцы яна заставалася знешне здаровай. У бярозы i асiны пры такой дозе апраменьвання анамалiі адсутнічалі. Пры паглынутых дозах 1,5–2,0 Гр дэфармацыi пупышак, iглiцы i парасткаў практычна не было.
Утрыманне хларафiлу. Адным з асноўных паказчыкаў фiзiялагiчнай актыўнасцi асiмiляцыйнага апарату раслiн з'яўляецца аптымальнае ўтрыманне хларафiлу. На гэты паказчык аказваюць уплыў лесагадоўчыя ўмовы, шматлiкiя фактары навакольнага асяроддзя, асаблiвасцi дрэвавых парод i iнш. У лiтаратуры сустракаюцца звесткi аб спрыяльным уплыве адносна малых (0,5–5 кР) i адмоўным больш высокiх доз iанiзуючага выпраменьвання на ўтрыманне хларафiлу ў лiсцi сельскагаспадарчых раслiн.
Разавае вострае апраменьванне iглiцы сасны ў дозе 20 Гр, па даных Панамаровай Р. П., прыводзiць да знiжэння ўтрымання пiгментаў пластыды ў 2–7 разоў i павелiчэння колькасцi хларапласту ў 1,7–2,3 раза.
Кудзiнавым М. А. адзначана знiжэнне ўтрымання хларафiлу у лiсцi аднагадовых сеянцаў ясеня арэгонскага i звычайнага, што вырашчаны з апрамененага насення на 18–40% у дозе 5–15 кР i на 69% у дозе 30 кР.
Даследаваннямi супрацоўнiкаў IЭБ НАНБ (Парфёнаў В. I. i iнш.) у зоне ЧАЭС устаноўлена, што пры забруджваннi глебы радыенуклiдамi да 100–220 Кi/км2 у iглiцы саджанцаў сасны назiраецца павышэнне ўтрымання хларафiлу на 10–20% у параўнаннi са слабаапрамененымi (кантроль). За межамi гэтага парога (да 350 Кi/км2) адбываецца паступовае знiжэнне ўтрымання хларафiлу да ўзроўню кантролю i толькi пры шчыльнасцi забруджвання звыш за 500 Кi/км2 велiчыня гэтага паказчыка ў розныя гады знiжалася на 11–15% у асноўным за кошт хларафiлу а. Пры павелiчэннi ў глебе ўтрымання радыенукiдаў назiраецца ўстойлiвае павышэнне ўтрымання жоўтых пiгментаў.
Вынiкi даследаванняў сведчаць, што знiжэнне ўтрымання хларафiлу ў лiсцях (iглiцы) шматлiкiх дрэвавых вiдаў да 50–60% не заўсёды адмоўна адбiваецца на iнтэнсiўнасцi iх росту.
Транспiрацыя. Даследаванне ўплыву iанiзуючых выпраменьванняў на iнтэнсiўнасць транспiрацыi дрэвывых вiдаў толькi пачынаецца. Вынiкi даследаванняў Кудзiнава М. А. з сеянцамi клёна ясенялiстага i дуба чырвонага, вырашчаных з апрамененага дозамi 5–30 i 5–10 кР насення, сведчаць аб прапарцыянальнай дозе апраменьвання, знiжэннi iнтэнсiўнасцi транспiрацыi. Пры сублятальнай дозе апошняя знiжалася на 50% i больш у параўнаннi з кантролем. Парушэнне воднага рэжыму прыводзiла да пагаршэння росту апрамененых раслiн. На гэтай падставе аўтар прыходзiць да высновы, што iнтэнсiўнасць транспiрацыi вельмi яскрава адлюстроўвае ступень прамянёвага ўздзеяння на раслiны.
Забруджванне глебы радыенуклiдамi ў зоне да 100–120 Кi/км2 не аказвае ўплыву на iнтэнсiўнасць транспiрацыi саджанцаў сасны (Парфёнаў В. I., Якушаў Б. I., Марцiновiч Б. С. i iнш). Пры павышэннi забруджанасцi глебы да 380 Кi/км2 i больш iнтэнсiўнасць транспiрацыi знiжаецца на 18–30%. Такая шчыльнасць забруджвання глебы адмоўна ўплывае на асноўныя параметры росту культур сасны.
Фотасінтэз. Уплыў iанiзуючага выпраменьвання на фотасiнтэз дрэвавых вiдаў даследаваны вельмi слаба. Пры разавым апраменьваннi iглiцы сасны ў дозе 10 Гр адзначана стымуляванне iнтэнсiўнасцi фотасiнтэзу ў 1,5 раза, у дозе 20 Гр адмоўнага ўплыву на гэты працэс не назiралася (Панамарова Р. П.). Была таксама адзначана затрымка росту i знiжэнне асiмiляцыйнай паверхнi iглiцы ў год апраменьвання.
Па даследаваннях Кудзiнава М. А., у двухгадовых сеянцаў ясеня арэгонскага, вырашчаных з апрамененага дозамi 0,5–15 кР насення, працэс фотасiнтэзу працякаў у 3–16 разоў актыўней, чым у кантрольных. У ясеня звычайнага на сёмым годзе пасля апраменьвання iнтэнсiўнасць гэтага працэсу перавышала кантрольную на 30–40% у залежнасцi ад дозы. Пры апраменьваннi сфармiраваных раслiн працэс фотасiнтэзу практычна не змяняўся.
Забруджванне глебы радыенуклiдамi ў зоне ЧАЭС у дозе 105 i 224 Кi/км2 не аказвала ўплыву на iнтэнсiўнасць фотасiнтэзу. Павышэнне шчыльнасцi забруджвання да 378 Кi/км2 садзейнiчала знiжэнню iнтэнсiўнасцi фотасiнтэзу на 12%. Павышенне дозы апраменьвання садзейчала далейшаму знiжэнню асiмiляцыi СО2 (Парфёнаў В. І. і інш.).
Рост i развiццё дрэў i дрэвастояў. Найбольш значныя радыебiялагiчныя эфекты ў дрэвавых пародах праяўляюцца ў першыя месяцы i гады пасля апраменьвання. У далейшым назiраецца паступовае знiжэнне адмоўных вынiкаў радыенуклiднага забруджвання i пераважанне аднаўленчых працэсаў. Ступень i характар пашкоджання дрэў залежыць ад дозавых нагрузак. Па даследаваннях Карабань Р. Т. i iншых пры дозе апраменьвання да 25 Гр прырост у кантрольных i апрамененых сосен быў блiзкiм, пры дозе 25–30 Гр ён знiжаўся, а пры дозе 40 Гр практычна адсутнiчаў. З павелiчэннем дозы апраменьвання змяншалася колькасць дрэў, якiя давалi прырост. Сублятальнымi паглынутымi дозамi для прыросту аўтары лiчаць 25–30 Гр. Пры больш высокiх назiралася ўсыханне дрэў. У дыяпазоне доз да 25 Гр знiжэнне прыросту абумоўлiвалася пашкоджаннем iглiцы i лiсця, вышэй за 25 Гр – пашкоджаннем пупышак i камбiю. У бярозы прыметнай рознiцы ў радыяльным прыросце ў кантрольных i апрамененых дрэў дозамi да 100 Гр у год апраменьвання не назiралася.
Па даследаваннях Козубава Г. М. i Таскаева А. I. пасля аварыi на ЧАЭС у зоне лятальнага пашкоджання (паглынутая доза 80–100 Гр) загiнулi практычна ўсе дрэвы, сублятальнага (паглынутая доза 8–25 Гр) – псавалiся часткова i пераважна ў маладняках i нiзкапаўнотных насаджэннях. У зоне сярэдняга пашкоджання (паглынутая доза 3–4 Гр) адзначана пагаршэнне роставых працэсаў, раздваенне радоў трахеiд, зубчатасць, скрыўленасць асяродачных прамянёў, моцныя ваганнi ў iх памерах, заўчаснае ападанне iглiцы, знiжэнне бягучага прыросту ў маладых дрэў з павышанай энергiяй росту, зрух працэсу нарастання гадавога кольца i павелiчэнне долi позняй драўнiны. У зоне слабага пашкоджання (паглынутая доза 0,5–1,0 Гр), якая займае асноўную частку 30-кiламетровай зоны i раёнаў, што прылягаюць да яе, выяўлены толькi храмасомныя анамалii ў меёзе i знiжэнне ўсходжасцi насення.
Знiжэнне паказчыкаў росту сеянцаў сасны з насення ўраджаю 1987 г., сабранага ў зоне ЧАЭС, знаходзiлася ў прамой залежнасцi ад узроўню забруджвання глебы (Парфёнаў В. І. i iнш.).
У год радыяцыйнага пашкоджання ў сасны адбывалася моцнае знiжэнне прыросту верхавiнкавых парасткаў. Пры гэтым найбольш значны ўплыў на іх рост назiраўся ў дрэў III–V класаў росту (па Крафту Г.).
Размеркаванне дрэў па катэгорыях санiтарнага стану. Усыханне i адмiранне дрэў пры ўздзеяннi iанiзуючага выпраменьвання з'яўляецца адным з самых важных iнтэгральных паказчыкаў змяненняў на бiягеацэнатычным узроўнi. Iндэкс пашкоджанасцi дрэвастояў таксама звязаны з узроўнямi тэхнагеннага радыяцыйнага забруджвання. Пры паглынутых сасной дозах вышэй за 100 Гр поўнае ўсыханне адбываецца праз тры месяцы пасля апраменьвання ў перыяд адноснага фiзiялагiчнага спакою. Дрэвы, на якія ўздзейнічала доза 70–100 Гр, усыхаюць да канца першага года пасля апраменьвання. Сублятальнай дозай (ЛД50) для сасны па крытэрыю ўсыхання дрэў лiчыцца 20–40 Гр, лятальнай 70–80 Гр (Карабань Р. Т. i iнш.). Лятальныя эфекты ў бярозы нават пры звышлятальных дозах адбывалiся толькi на другi i трэцi год. Поўнае ўсыханне пачалося на другi год пасля апраменьвання. Лятальная доза (ЛД100) незалежна ад класа росту дрэў склала 600–800 Гр. На трэцi год ЛД100 складала 250 Гр. Да канца трэцяга года дрэвы III класа росту ўсыхалi пры дозах 150–200 Гр.
Пашкоджанне насаджэнняў, выклiканае апраменьваннем, суправаджалася змяншэннем супрацiўляльнасцi ў адносiнах да насякомых, грыбоў i бактэрый. Узмоцненае размнажэнне энтама-шкоднiкаў у аслабленых апраменьваннем дрэвастоях з'яўляецца тыповым другасным радыяцыйным эфектам у лясах. Пашкоджанне насаджэнняў у вынiку масавага размнажэння шкоднiкаў можа быць больш моцным, чым ад непасрэднага ўздзеяння радыяцыi.
12.7.6. Уплыў iанiзуючага выпраменьвання на рэпрадуктыўную сферу фiтацэнозаў
Праведзеныя ў пачатку 60-х г. XX ст. і працягнутыя ў далейшым даследаваннi па ўплыве g-выпраменьвання на розныя вiды сосен паказалi, што найбольш адчувальнай да ўздзеяння iанiзуючага выпраменьвання з'яўляецца iх репрадуктыўная сфера, ад фармiравання i стану якой залежыць якасць насення. Нават пры слабых дозах апраменьвання пашкоджваецца структура рэпрадуктыўных органаў, што прыводзiць да знiжэння паўназярнiстасцi i ўсходжасцi насення (Козубаў Г. М. i iнш.). Паслядзеянне апраменьвання, якое назiралася на працягу 10 гадоў i больш, выклiкала стэрыльнасць апрамененых раслiн i бясплоднасць дачынных арганiзмаў (Цiхамiраў Ф. А., Аляксахiн Р. М., Кудзiнаў М. А.). Сублятальныя дозы выклiкаюць амаль поўную гiбель рэпрадуктыўных тканак. Пасля апраменьвання сасны цвёрдаiглiчнай у дозе 2,2 Р/дзень (Spаrrow А. І., Woodwell) назiралася павелiчэнне аб'ёму ядра, рэдукцыі колькасцi храмасом, падаўжэнне термiну меёзу, храмасомныя анамалii i iншыя парушэннi генетычнага цыклу. Пасля хранiчнага апраменьвання мужчынскiх пупышак восенню колькасць храмасомных аберацый узрастала з павелiчэннем магутнасцi дозы ад 1% пры 0,4 Р/дзень да 76% пры 17 Р/дзень. Адпаведна знiжалася апладняльная здольнасць пылку. Памеры мiкрастрабiлаў пачыналi змяншацца пры дозе 9 Р/дзень i больш. Пры 56 Р/дзень меёз у мiкраспорацытах затрымлiваўся на 17 дзён. Пры апраменьваннi мужчынскiх пупышак сасны звычайнай у дозах ад 6 да 22 Гр паспяванне пылку затрымлiвалася на 2–3 днi (Фёдараў Е. А., Цiхамiраў Ф. А.). Вострае i хранiчнае апраменьванне пры павышаных дозах прыводзiла да поўнай гiбелi як жаночых, так i мужчынскiх парасткаў (Козубаў Г. М., Таскаеў А. І.) на участках з сублятальным i сярэднiм пашкоджаннем пры паглынутых дозах па g-выпраменьваннi 2 Гр i больш. У першы вегетацыйны перыяд на кантрольных участках i участках са слабым узроўнем апраменьвання закладка новых мужчынскiх пупышак працякала нармальна, з сярэднiм – толькi ў асобных дрэў. Самая вялiкая колькасць стэрыльнага пылку ўтвараецца пры зыходнай экспазiцыйнай дозе 40–50 мР/гадз., самая малая – пры 20–25 мР/гадз. У цэлым уплыў iанiзуючага выпраменьвання на мужчынскую рэпрадуктыўную сферу сасны звычайнай адбiваецца практычна на ўсiх стадыях мiкраспорагенезу i фармiравання мужчынскага гаметафiту i праяўляецца як у змяненнi тэрмiнаў наступлення мiкрафеналагiчных фаз, так i ў знiжэннi жыццяздольнасцi i фiзiялагiчнай актыўнасцi пылку (Козубаў Г. М., Таскаеў А. І.).
Парушэннi ў ходзе меёзу ў семяпупышках сасны звычайнай пры адносна слабым iанiзуючым уздзеяннi не аказваюць iстотнага ўплыву на далейшае развiццё жаночага гаметафiту i толькi ў некаторых выпадках анамалii ў мета- i анафазе першага i другога дзяленняў меёзу знiжаюць колькасць семяпупышак з нармальна развiтым жаночым гаметафiтам. Пры паглынутых дозах 2–4 Гр назiралiся значныя парушэннi на працягу ўсяго мiкраспорагенезу; 0,7–1,1 Гр – колькасць некрацiраваных насенных пупышак не перавышала 5%; 1,7–2,3 Гр – 50%; 3,5–4,7 Гр – 70%.
Шышкi другога года развiцця пасля вострага i хранiчнага апраменьвання (Козубаў Г. М., Таскаеў А. І.) пры дозе 800–1000 Гр знешне не адрознiвалiся ад неапрамененых i дасягалi нармальных памераў i масы. Бiялагiчныя ўласцiвасцi насення i яго колькасныя паказчыкi пацвердзiлi выразную залежнасць ад узроўню радыяцыйнага ўздзеяння. Пры апраменьваннi ў дозе 8–10 Гр усходжасць насення складала ўсяго 3%. Знiжэнне яе адбывалася, у асноўным, за кошт павелiчэння колькасцi пустога насення i некрозу жаночага гаметафiту ў перыяд вострага апраменьвання. Сувязi пасяўных якасцей насення наступных за апраменьваннем гадоў не назiралася.
Найбольш выразны ўплыў iанiзуючага выпраменьвання на бiялагiчныя ўласцiвасцi насення адзначаўся пасля вострага апраменьвання дозай звыш за 8–10 Гр, якое прыводзiла практычна да поўнай гiбелi жаночага гаметафiту на стадыi свабодных ядраў i 100% утварэння пустога насення. Спелае насенне даволi ўстойлiвае да апраменьвання. Яно здольнае вытрымлiваць больш высокiя дозы, чым мацярынскiя дрэвы (Кудзiнаў М. А.). Гэта звязана з тым, што клеткi зародыша маюць больш дробныя храмасомы ў параўнаннi з храмасомамi саматычных клетак раслiн, што вегеціруюць. Крытычныя дозы апраменьвання насення сасны, елкi, пiхты, лiстоўнiцы i iншых хваёвых, якiя выклiкаюць знiжэнне ўсходжасцi на 50% i далейшую гiбель значнай часткi ўсходаў, складаюць пры вострым апраменьваннi ад 6 да 60 Гр; лiставых парод – больш за 100 Гр (табл. 12.6).
Таблiца 12.6
Дозы вострага g-выпраменьвання, якiя знiжаюць усходжасць насення асноўных лесаўтваральных парод на 50% (Цiхамiраў Ф. А., Кудзiнаў М. А.)
| Парода | Крытычная паглынутая доза, Гр |
| Сасна веймутава | |
| Сасна звычайная | >30<55; 25; >7<15; 50 |
| Сасна Банкса | 63; 49 |
| Елка звычайная | 8–30; 40; >7<15 |
| Лiстоўнiца сiбiрская | >10<50; 75; >7<15 |
| Кедр сiбiрскi | >10<50 |
| Пiхта сiбiрская | >50<100 |
| Вольха чорная | |
| Бяроза бародаўчатая | >50<100 |
| Бяроза пушыстая | 100; >50<100 |
| Асiна звычайная | |
| Дуб чарэшчаты | |
| Клён востралiсты | |
| Клён ясенялiсты | |
| Ясень звычайны | |
| Лiпа буйналiстая | |
| Акацыя жоўтая | >150<100 |
| Акацыя белая |
Устойлiвасць насення да апраменьвання, на думку Кудзiнава М. А., фармiравалася ў працэсе эвалюцыi не як спецыфiчная рэакцыя арганiзма на iанiзуючае выпраменьванне, а як унiверсальная рэакцыя на неспрыяльныя фактары навакольнага асяроддзя, у асноўным на дэфiцыт глебавай i паветранай вiльгацi. Павышаная рэзiстэнтнасць цесна карэлюе са здольнасцю раслiн адаптавацца да неспрыяльных умоў навакольнага асяроддзя, у тым лiку і да засухi. Чым шырэйшы арэал распаўсюджвання вiду, тым вышэйшая i радыеадчувальнасць.
У залежнасцi ад вiльготнасцi насення, тэрмiнаў i лесагадоўчых умоў мацярынскiх дрэў лятальныя дозы апраменьвання адных i тых жа вiдаў яго вагаюцца ў шырокiм дыяпазоне.
У прыродных умовах устойлiвасць насення да апраменьвання i выжывальнасць усходаў звычайна значна нiжэйшыя, чым у лабараторных (Карабань Р. Г., Кудзiнаў М. А.). Лятальныя дозы ў прыродных умовах пасоўваюцца ў бок больш нiзкiх значэнняў. Вынiкi апраменьвання ў некаторым скарачэннi рэпрадуктыўнай здольнасцi праяўляюцца нават у наступным пакаленнi.
Рэакцыя насення на апраменьванне не абмяжоўваецца толькi паталагiчнымi змяненнямi. Пры адносна невялiкiх дозах (да 10–20% ад лятальных значэнняў) магчыма стымуляцыя iх грунтовай усходжасцi на 15% i больш i ўзмацненне росту сеянцаў. Радыестымуляцыя ў некаторых вiдаў сасны, елкi i лiстоўнiцы выяўлена Карабань Р. Т., а для лiставых парод Кудзiнавым М. А.
Пры дастакова высокiх узроўнях радыеактыўнага забруджвання апраменьванне можа выклiкаць гiбель не толькi ўсяго дрэвавага яруса, але i глебавага запасу насення i прывесцi да пагаршэння або спынення насеннага ўзнаўлення. Грунтовая ўсходжасць насення елкi звычайнай пры паглынутай дозе каля 6 Гр знiжалася на 67%, сасны i лiстоўнiцы – на 20% (Крывалуцкi Д. А.). Пры паглынутай дозе каля 20 Гр усходжасць насення поўнасцю страчвалася.
Да канца першага вегетацыйнага перыяду сеянцы на ўчастках, дзе паглынутая доза ў тканках іх апiкальнай мерыстэмы складала каля 20 Гр, раслiны па знешнім выглядзе моцна адрознiвалiся ад канрольных. Iглiца ў нiжняй частцы сеянцаў жаўцела i ўсыхала, кроны былi ажурныя, назiралася недаразвiтасць або адмiранне бакавых пупышак, а часам больш моцнае iх развiццё, знiжэнне паказчыкаў росту на 20–25% (Крывалуцкi Д. А. i iнш.). Пры меншых узроўнях забруджвання (паглынутая доза за 3 месяцы да 6 Гр) сеянцы па развiцці не адрознiвалiся ад кантрольных. У наступны вегетацыйны перыяд паглынутая доза ў тканках апiкальнай мерыстэмы сеянцаў павялiчвалася да 84 Гр i ў вынiку адбывалася знiжэнне прыросту бiямасы, адставанне ў росце ў 2–3 разы, а таксама адмiранне верхавiнкавых пупышак у 90% сеянцаў. Аналагiчныя вынiкi атрыманы i праз пяць гадоў пасля апраменьвання, аднак эфекты былi больш слабымi. Адпад сеянцаў у сасны складаў 62%, лiстоўнiцы – 71%, елкi – 55%, на кантрольных адпаведна 21, 15 i 13%. Сеянцы, што захавалiся да канца другога вегетацыйнага перыяду, былi слабымi i нежыццяздольнымi. Прыжывальнасць сеянцаў i саджанцаў пры штучным насенным аднаўленнi таксама залежыць ад дозы iанiзуючага апраменьвання.
Механiзм вегетатыўнага аднаўлення бiягеацэнозаў у многiм вызначаецца бiялагiчнымi асаблiвасцямi дрэвавых парод i ўплывам iанiзуючага выпраменьвання на рэпрадуктыўную здольнасць раслiн. У асiны i бярозы пры дозах апраменьвання, якiя выклiкаюць усыханне крон, аднаўленне адбываецца за кошт з'яўлення ствалавых або каранёвых парасткаў. Праз два гады пасля радыяцыйнага пашкоджання надземнай часткi бярозы ад пнёў 60% загiнуўшых дрэў з'яўляюцца густыя цалкам жыццяздольныя парасткi, якiя ў наступныя гады нармальна развiваюцца. У асiны яны вырастаюць ад тонкiх каранёў. Здольнасць да вегетатыўнага аднаўлення ў гэтых парод праяўляецца нават пры апраменьваннi надземнай часткi дрэў зверхлятальнымi дозамi. Пры дозах менш за 300 Гр парасткi з'яўляюцца непасрэдна пасля апраменьвання. Больш высокiя дозы выклiкаюць затрымку іх з'яўлення да двух гадоў. Аднаўленне лiставых насаджэнняў, пашкоджаных у вынiку апраменьвання, залежыць ад узросту дрэў, iх фiзiялагiчнага стану, лесагадоўчых умоў i iншых фактараў.
Пры паглынутых дозах 250–800 Гр стваловыя парасткi з'яўляюцца толькi ў 10% пашкоджаных дрэў (Карабань Р. Т.), пры 15–20 Гр – у 50–60%. Першапачаткова парасткi з'яўляюцца ў дрэў нiжэйшых класаў росту, у больш маладых у горшых лесагадоўчых умовах. Паглынутыя дозы ў 250–300 Гр не толькi выклiкаюць гiбель бярозы, але i практычна цалкам заглушаюць аднаўляльную здольнасць дрэў усiх класаў росту.
Розная радыеадчувальнасць вiдаў у зонах радыеактыўнага забруджвання прыводзiць да абяднення вiдавой разнастайнасцi, змянення рэпрадуктыўных, роставых i iншых працэсаў, знiжэння прадукцыйнасцi i ўстойлiвасцi лясных бiягеацэнозаў.
У вынiку катастрофы на ЧАЭС у зоне радыяцыйнага забруджвання аказалiся аб'екты каштоўнага селекцыйнага фонду, якiя ўваходзiлi ў пастаянную лесанасенную базу рэспублiкi. Гэта лясныя генетычныя рэзерваты, плюсавыя насаджэннi i дрэвы, клонавыя насенныя плантацыi i лесанасенныя ўчасткi. Значная частка гэтых аб'ектаў, акрамя вострага апраменьвання 1986 г., адчувае на сабе тыя цi iншыя дозы хранiчнага апраменьвання. Па гэтай прычыне паўстала пытанне аб неабходнасцi вызначэння характару i ступенi пашкоджання спадчыннага апарату, зыходных бацькоўскiх фенатыпаў i мэтазгоднасцi выкарыстання дадзеных участкаў у лесанасеннай гаспадарцы для арганiзацыi нарыхтоўкi насеннай прадукцыi для лесааднаўлення (Iпацьеў В. А. i iнш.). Аўтары назiралi выразнае павелiчэнне назапашвання радыецэзiю ў шышках сасны з узрастаннем радыяцыйнага забруджвання глебы. Пры шчыльнасцi яе забруджвання 370–1480 кБк/м2 удзельная радыеактыўнасць шышак у сярэднiм не перавышала 3000 Бк/кг, пры 2590–3700 кБк/м2 вагалася ў межах 4440–16 280 Бк/кг, 3700 - 5500 кБк/м2 перавышала 7500 Бк/кг, а пры шчыльнасцi забруджвання глебы звыш за 9250 кБк/м2 яна дасягала 18 000–133 000 Бк/кг.
Выхад насення з насаджэнняў 30-кiламетровай зоны ў сярэднiм знiжаўся ў 1,5 раза. Дрэвы, якiя пацярпелi ад вострага апраменьвання, мелi больш дробныя шышкi. Велiчыня i маса iх з непашкоджаных апраменьваннем дрэў былi блiзкiмi да кантрольных.
Хранiчнае апраменьванне дрэў у розных зонах радыяцыйнага забруджвання i нават на ўчастках лятальнага пашкоджання насаджэнняў стымулявала энергiю прарастання i лабараторную ўсходжасць насення. Значных адрозненняў у даўжынi праросткаў насення сасны з хранiчна апраменьваемых эксперыментальных участкаў зон слабага, сярэдняга i сублятальнага пашкоджання не назiралася. З насення дрэў зоны лятальнага пашкоджання фармiравалiся менш развiтыя праросткi. Усходжасць насення з хранiчна апрамененых насаджэнняў у гадавальнiках была на 14–28% нiжэйшая за кантрольную. Адрозненняў у захаванасцi сеянцаў з насення кантрольных i хранiчна апраменьваемых насаджэнняў у канцы першага вегетацыйнага перыяду не назiралася. Пасля перазiмоўкi адпад сеянцаў з насення хранiчна апраменьваемых насаджэнняў быў вышэй за кантрольныя.
12.7.7. Уплыў iанiзуючага выпраменьвання на жывое наглебавае покрыва
· У лясных бiягеацэнозах жывое наглебавае покрыва з'яўляецца адным з фактараў, што фармiруюць пажыўнае асяроддзе. Радыеадчувальнасць яго на парадак вышэй, чым дрэвавага яруса. Лятальная доза для мхоў i лiшайнікаў вагаецца ў межах 2–5 тыс. Гр (Козубаў Г. М., Таскаеў А.І., Градзiнскi Д. М.). Для значнай часткi вiдаў травянiстых раслiн пад полагам апрамененага лесу магутнасць экспазiцыйнай дозы вышэй за 0,45 Гр/сут з'яўляецца лятальнай.
Прамянёвыя эфекты ў травянiстых раслiн пры апраменьваннi дозамi да 230 Гр нязначныя i выражаюцца ў кароткатэрмiновым пагаршэннi або паляпшэннi росту, прамянёвых апалах i марфалагiчных метамарфозах (Спiрын Д. А. i iнш.). На ўчастках са шчыльнасцю забруджвання звыш за 1500 Кі/км2 на працягу 3–4 гадоў пасля апраменьвання шматгадовыя раслiны поўнасцю гiнуць (Спiрын Д. А. i iнш.). Праз 2–3 гады пасля завяршэння працэсу дэфалiяцыi пашкоджаных дрэў, ападзення галля i вывальвання загiнуўшых дрэў назiраецца паляпшэнне росту i развiцця травянога покрыва, назапашванне надземнай фiтамасы ў 1,5–3,5 раза ў параўнаннi з неапрамененымi насаджэннямi. Адбываецца змяненне вiдавога саставу за кошт вегетатыўнага размнажэння светлалюбiвых вiдаў (пажарніца, асокi, сунiчнiк) i выпадзення з саставу радыеадчувальных вiдаў (Цiхамiраў Ф. А.).
Розныя вiды травянiстых раслiн характарызуюцца неаднолькавым назапашваннем радыенуклiдаў. Ва ўмовах сасняку мшыстага па ступенi назапашвання γ-утрымлiваючых радыенуклiдаў травяна-кустовую раслiннасць размяркоўваюць у наступным парадку: iмхi > папаратнiкавыя > залознiкавыя (мар'яннiкi) > драсенавыя > бруснiчныя (бруснiчнiк, чарнiчнiк) > святаяннiкавыя складанакветкавыя > лiлейныя (ландыш > купена лекавая, майнiк двухлicты) > губакветныя (чабор) > ружакветныя (сунiчнiк) > злакавыя > верасовыя > грушанкавыя (Парфёнаў В. I. i iнш.). Рознiца ў канцэнтрацыi 90Sr i 137Cs, адзначаная ў розных вiдаў раслiн i iх частках, можа перавышаць 10-кратную велiчыню. Найбольш высокiя канцэнтрацыi 90Sr i 137Cs адзначаны ў фотасiнтэзуючых частках раслiн. Імхова-лiшайнiкавая раслiннасць назапашвае радыенуклiды ў значна большых колькасцях, чым шматлiкiя вiды травянiстых раслiн. Роля мхоў як акумулятараў радыенуклiдаў вызначаецца iх водаўтрымлiваючай здольнасцю. Высокая самкнутасць iмховага покрыва стварае ўмовы для працяглага ўтрымання ў iм водных раствораў i назапашвання радыеактыўных рэчываў, якiя паступаюць з атмасферы i паверхневых сцёкаў. У перыяд iнтэнсiўных выпадзенняў ападкаў iмховае покрыва пры суцельным пакрыццi можа затрымаць да 100% 90Sr, якi паступае з атмасферы.
Усе вiды лекавых раслiн, якiя растуць у гiдраморфных умовах, назапашваюць у дзесяткi разоў большую колькасць 137Cs, чым у аўтаморфных. У сувязi з гэтым лекавыя раслiны-гiграфiты характарызуюцца максiмальным забруджваннем. Пры павышаным увiльгатненнi глебы назапашванне 137Cs ягадамi ўзрастае ў 17 разоў i па ступенi забруджвання значна пераўзыходзiць лiсце.
На назапашванне радыенуклiдаў у лекавых раслiнах лясных фiтацэнозаў значны ўплыў аказваюць лесагадоўчыя ўмовы i вiдавая прыналежнасць. Ландыш майскi, купена лекавая i буквiца лекавая ва ўмовах дубровы арляковай назапашваюць радыенуклiдаў у 5–6 разоў больш, чым бруснiчнiк i чарнiчнiк (Парфёнаў В. I. i iнш.).
Вынас радыенуклiдаў у фiтамасу раслiн адбываецца ў асноўным з пласта глебы да 30 см. Для вiдаў з паверхневай каранёвай сiстэмай (канюшына паўзучая, дуброўка, гарошак мышыны) зонамi найвялiкшага паглынання з'яўляюцца пласты 0–5 i 0–10 см. Канюшына лугавая, ракiтнiк рускi i дрок фарбавальны паглынаюць 90Sr з пласта 0–30 см больш – менш раўнамерна. Вынас радыенуклiдаў з розных пластоў глебы знаходзiцца ў прамой залежнасцi ад вiльготнасцi, уласцiвасцей глебы i бiямасы актыўных каранёў. Ва ўмовах беларускага Палесся, дзе пераважаюць дзярнова-падзолiстыя i тарфяныя глебы з нiзкай ёмiстасцю паглынання, слабай насычанасцю калiем, кальцыем i магнiем, нiзкiм утрыманнем рухомых формаў мiкраэлементаў, назiраецца павышанае назапашванне радыенуклiдаў у наглебавым покрыве.
Назапашванне радыенклiдаў падлескам таксама залежыць ад узроўню забруджвання iмi глебы, вiдавой прыналежнасцi i лесагадоўчых умоў.
12.7.8. Мiграцыя радыенуклiдаў у глебе
Глеба – адно з важнейшых звёнаў мiграцыi радыенуклiдаў у прыродных ландшафтах. Радыеактыўныя рэчывы, якiя паступаюць у атмасферу ў вынiку аварый, ядзерных выпрабаванняў, работы прадпрыемстваў атамнай прамысловасцi i з iншых крынiц, паступова асядаюць на наглебавае покрыва, сарбiруюцца глебай i становяцца даступнымi для засваення каранёвымi сiстэмамi раслiн. Паступленне iх з атмасферы i замацаванне ў подсцiле i верхнiм гарызонце глебы забяспечвае iснаванне працягла дзеючай крынiцы радыенуклiдаў для раслiн. Моцная сорбцыя радыенуклiдаў цвёрдай фазай глебы i магчымы пераход з абменнай формы ў неабменную абмяжоўваюць iх мiграцыю i засваенне раслiнамi праз каранёвыя сiстэмы. Каранёвае паглынанне радыенуклiдаў раслiнамi залежыць яшчэ i ад уласцiвасцей глебы i глебавай хiмii радыенуклiдаў. Для ацэнкi паступлення радыенуклiдаў з глебы ў раслiны выкарыстоўваюць розныя паказчыкi (каэфiцыент назапашвання, каэфiцыент прапарцыянальнасцi i iнш.).
Хуткасць пераносу мiкраколькасцi радыенуклiдаў па трафiчных ланцугах у сiстэме «глеба–раслiны–глеба» залежыць ад утрымання iзатопных (стабiльны 88Sr, радыеактыўны 90Sr) або неiзатопных (90Sr i стабiльны 40Ca) аналагаў. Сярод фiзiка-хiмiчных характарыстык глебы самы вялiкi ўплыў на паводзiны радыенуклiдаў i пераход iх у раслiны аказвае ўтрыманне абменных катыёнаў, арганiчнага рэчыва, фiзiчнай глiны i глею, ёмiстасць паглынання, механiчны састаў i вiльготнасць глебы. Гэта абумоўлiвае своеасаблiвыя заканамернасцi мiграцыi радыенуклiдаў у лясных бiягеацэнозах.
У першыя днi пасля катастрофы на ЧАЭС каля 70–80% усiх радыеактыўных рэчываў было затрымана надземнымi часткамi дрэвавых парод i каля 20–30% – наглебавым покрывам (Якушаў Б. І., Марцiновiч Б. С., Цiхамiраў Ф. А. i iнш.). Прычым сасновыя насаджэннi затрымлiвалi ў 2–3 разы больш лiставых i ў 7–10 разоў больш за iншыя прыродныя цэнозы (лугавыя, балотныя). У далейшым адбывалася пераразмеркаванне радыенуклiдаў па кампанентах бiягеацэнозаў. Перыяд паўачышчэння крон складаў 160–170 дзён (Якушаў Б. І.). У залежнасцi ад тыпу фiтацэнозу (Цiхамiраў Ф. А.) утрыманне радыенуклiдаў у дрэвавым ярусе складала 6–7% ад першапачатковага. Асноўная колькасць iх перамяшчалася ў лясны подсцiл, а потым – у мiнеральныя пласты глебы.
У цяперашнi час радыяцыйнае становiшча вызначаецца доўгажывучымi нуклiдамi 90Sr, 137Cs, 238Pu, 239Pu, 240Pu. У глебе радыенуклiды знаходзяцца ў чатырох формах: воднарастваральнай, абменнай, кіслотнарастваральнай i нерухомай. Раслiны паглынаюць з глебы рухомыя формы воднарастваральных радыенуклiдаў, якiя знаходзяцца ў зоне распаўсюджвання iх каранёвых сiстэм. Колькасць радыенуклiдаў, бiялагiчныя асаблiвасцi раслiн i доля рухомых формаў вызначаюць ступень назапашвання iх у раслiнах. Каэфiцыенты мiграцыi радыеактыўных 90Sr i 137Cs залежаць ад механiчнага саставу глебы. 137Cs хутчэй фiксуецца глебай i таму яго даступнасць для раслiн нiжэйшая, чым стронцыю. Утрыманне ў глебе яго рухомых формаў з цягам часу практычна не змяняецца i складае 76–90% ад агульнай яго колькасцi ў верхнiм пласце глебы (Раманаў Д. А., Спiрын Д. А., Аляксахiн Р. М.).
Аненкаў Б. М. i Юдзiнцава К. В. лiчаць, што 90Sr паступае ў надземную частку дрэвавых раслiн з глебы ў значна большай колькасцi, чым 137Cs. Сярод дрэвавых парод сасна звычайная i лiстоўніца сібірская характарызуюцца ў 2–7 разоў меншым у параўнаннi з елкай назапашваннем 90Sr. Парфёнавым В. I. i iншымі устаноўлена, што ў зоне ЧАЭС ва ўсiх тыпах лесу, у раслiнах i глебе ўтрыманне 137Cs значна большае, чым 90Sr.
Пераразмеркаванне 137Cs у сасновых насаджэннях на глебах з лёгкiм механiчным саставам пры шчыльнасцi забруджвання 13 Кi/км2 за 1989–1990 гг. адбывалася толькi ў 5-сантыметровым пласце (Iпацьеў В. А. i iнш.). З павелiчэннем шчыльнасцi забруджвання да 100 Кi/км2 значная яго колькасць перамяшчалася ў пласт 10–15 см, а звыш за 400 Кi/км2 – на глыбiню 15–20 см. У насаджэннях з несфармiраваўшымся подсцiлам мiграцыя радыецэзiю па профiлi глебы адбывалася больш хутка. Праз пяць гадоў пасля катастрофы ўтрыманне яго ў подсцiле масай 7 т/га складала 21%, 20 т/га –48% i 40 т/га – 62%, у подсцiле i 5-сантыметровым пласце глебы – 94–97% ад агульнага запасу iх у глебе. Для 90Sr характэрная такая ж заканамернасць, як i для 137Cs, але мiграцыя яго працякае значна хутчэй.
Мiграцыя радыенуклiдаў па глебавым профiлi i назапашванне iх раслiнамi залежыць ад формы iх знаходжання ў глебе. На аўтаморфных глебах асноўная маса 137Cs знаходзiцца ў «фiксаванай» форме, водарастваральная форма яго складае 1,8–8%, абменная – 1,3–35%, рухомая – 5,3–56,1%. На гiдраморфных глебах доля абменных i рухомых формаў 137Cs больш высокая, чым на аўтаморфных.
Асноўная маса 90Sr у глебе знаходзiцца ў рухомай форме i складае 65,5–98,5% ад агульнага яго ўтрымання ў верхнiм 5–6-сантыметровым пласце, у тым лiку водарастваральная – 0,7–11,4%. Утрыманне 90Sr у «фiксаванай» форме вагаецца ў межах 1,7–34,5%.
Праз сем гадоў пасля катастрофы на ЧАЭС асноўная маса радыенуклiдаў сканцэнтравалася ў наглебавым покрыве i пяцiсантыметровым пласце глебы (Градзiнскi Д. М.).
У жывым наглебавым покрыве, лясным подсцiле i ў 5-сантыметровым пласце глебы акумулявана ад 70 да 97% радыенуклiдаў. На глыбiнi 5–10 см утрымлiваецца ад 2 да 19% радыенуклiдаў, 10–15 см – 1–8% i на глыбiнi 15–20 см – да 7%. Пры гэтым глыбiня i скорасць мiграцыi радыенуклiдаў залежаць у асноўным ад тыпаў лесу, фiзiка- i аграхiмiчных уласцiвасцяў i воднага рэжыму глебы, фiзiка-хiмiчных уласцiвасцей выпаўшых радыенуклiдаў.
137Cs сарбiруецца глебай i ў дадзены момант 70–83% яго знаходзiцца ў «фiксаваным» стане i таму мае нiзкую хуткасць вертыкальнай мiграцыi. 90Sr замацоўваецца па тыпе iоннага абмену i 92–95% запасу яго знаходзiцца ў рухомай форме.
Праз 33 гады пасля ядзернай аварыi на Паўднёвым Урале ў зоне радыеактыўнага следу шчыльнасць забруджвання глебы 90Sr перавышала кантроль у 7–50 разоў (Ааркрог А. i iнш.). Максiмальная канцэнтрацыя 90Sr (600 Бк/кг) адзначана ў пяцiсантыметровым пласце глебы. Каля 75% 90Sr i 90% 137Cs ад агульнага запасу iх у глебе сканцэнтравана ў пласце да 10 см.
Праз чатыры гады пасля ўнясення 90Sr i 137Cs у лясны подсцiл абодва радыенуклiды мiгрыруюць у гарызантальным кiрунку (Кулiкова i iнш.). Асноўная доля радыенуклiдаў застаецца ў зоне першапачатковага ўнясення. Найбольшай мiграцыйнай здольнасцю ва ўсiх тыпах лясных бiягеацэнозаў характарызуецца стронцый-90. Хуткасць дыфузii 90Sr і 137Cs складае адпаведна 0,8 і 0,5 см/год. З павышэннем увiльгатнення дзярнова-падзолiстых, дзярнова-лугавых i тарфяна-глеевых глеб (ад узроўню вiльготнасцi завядання да поўнай вiльгацяёмiстасцi) агульны вынас 90Sr, 137Cs i 144Се павялiчваецца.
Для паскарэння ачышчэння лясных земель ад дозаўстваральных радыенуклідаў супрацоўнікамі ІЛ НАН Беларусі Іпацевым В. А., Булко М. І. і іншымі распрацаваны спецыяльны антынуклідны глебава-раслінны бар’ер «біялагічная перагародка». Аўтары лічаць, што за кошт накіроўваемага рэгулявання ўрадлівасці глебы і ўтрымання ў лясным ападзе дозаўтваральных радыенуклідаў магчыма грунтоўна зніжаць іх назапашванне, вертыкальны рух па глебавым профілі і забяспечваць рэгулюемую ў рызасферы сінхронную міграцыю радыенукліда 137Cs у глыбіню глебы.
Па меркаванню Булавіка І. М. і Перавалоцкага А. Н. выкарыстанне мінеральных угнаенняў і пабудовы біялагічнай перагародкі не эфектыўна пры правядзенні рэабілітацыі лясоў забруджаных радыенуклідамі. Гэта звязана з асаблівасцямі характэрнымі толькі лясным экасістэмам і дакладнасцю выяўлення заканамернасцей паводзін радыенуклідаў у розных лесагадоўчых умовах. Неабходны пошук новых метадычных падыходаў і сумесных комплексных даследванняў на адзінай узгодненай метадычнай аснове.
Размеркаванне паглынутых доз b-выпраменьвання па вертыкалi ў глебе амаль iдэнтычнае размеркаванню ў профiлi глебы самiх радыенуклiдаў. У першыя месяцы пасля паступлення радыенуклiдаў iнтэнсiўнае апраменьванне зведваюць толькi насельнiкi ляснога подсцiлу. Паглынутая доза ў ім пры ўнясеннi 90Sr у колькасцi 106 Бк/м2 за першы год склала 0,3 Гр, праз 8 гадоў яна знiзiлася да 0,1 Гр у год. На працягу 13 гадоў радыенуклiды мiгрыравалi па профiлi на глыбiню да 5–9 см i аказвалi ўплыў на папуляцыi арганiзмаў, якiя насялялi гэты слой глебы (Крывалуцкi Д. А. i iнш.).
Аналiз колькасных паказчыкаў па мiграцыi 90Sr у розных звёнах транспарту радыенуклiдаў паказвае, што ў сасновых лясах 90Sr перамяшчаецца з подсцiлу ў глебу прыкладна ў 2,5–3 разы павольней, чым у бярозавых (каэфiцыент пераносу адпаведна роўны 0,17 i 0,47), што звязана з больш iнтэнсiўнай мiнералiзацыяй подсцiлу (Аляксахiн Р. М., Нарышкiн М. А.). З глебы ў бярозу 90Sr паступае прыблiзна ў шэсць разоў хутчэй, чым у сасну. 137Cs характарызуецца значна большай мабiльнасцю. З цягам часу асноўным месцам лакалiзацыi яго ў бiягеацэнозе становiцца глеба.
Моцны ўплыў на лясныя бiягеацэнозы аказвае плутонiй. Яго ўтрыманне ў глебах вагаецца ў межах 1–1100 Бк/кг, а ў раслiнах – 481–51 800 Бк/кг (Кабашнiкава).
Сабраныя ў апошнiя гады матэрыялы па ўтрыманнi i мiграцыi радыенуклiдаў у сiстэме «глеба–раслiна–глеба» даюць магчымасць будаваць мадэлi транспарту радыенуклiдаў. Наяўнасць такiх мадэляў дазволiць ацанiць уплыў асобных кампанентаў лясных бiягеацэнозаў у пераносе радыенуклiдаў.
12.7.9. Назапашванне радыенуклiдаў i ўздзеянне iанiзуючага выпраменьвання на лясную фауну
Лясная фауна з'яўляецца важным кампанентам прыродных i трансфармаваных экасiстэм. Шматлiкiя вiды служаць аб'ектам промыслу, крынiцай харчовых прадуктаў для чалавека, сыравiнай для вытворчасцi прамысловых тавараў i лекавых прэпаратаў. Таму ацэнка назапашвання i характару мiграцыi радыенуклiдаў у трафiчных ланцугах розных груп жывёлы, а таксама магчымых экалагiчных вынiкаў, мае вялiкае значэнне для распрацоўкi рэкамендацый па знiжэннi эфектаў радыяцыйнага забруджвання прыродных комплексаў.
Пры ацэнцы ўплыву радыяцыi на стан фауны, ў першую чаргу, неабходна ўлiчваць вiдавую радыеадчувальнасць. У якасцi бiялагiчнага параметра пры вызначэннi радыеадчувальнасцi на ўзроўнi арганiзма звычайна выкарыстоўваецца лятальная доза, якая выклiкае гiбель 50 або 100% асобiн за 30 сутак (ЛД50/30 або ЛД100/30). Для найбольш распаўсюджаных вiдаў у лясах прыблiзная сярэднелятальная доза выпраменьвання ЛД50/30 Гр складае
| Беспазваночныя | |||
| Амёбы | 1000–6000 | Членiстаногiя | 50–900 |
| Iнфузорыi | 3000–7000 | Асцыдыi | |
| Кiшэчнаполасцевыя | 50–2500 | Нематоды | 50–1000 |
| Малюскi | 120–200 | ||
| Насякомыя | |||
| Дарослыя | 800–2000 | Лiчынкi | 1–250 |
| Пазваночныя | |||
| Паўзуны | 80–200 | Рыбы | 5–20 |
| Чарапахi | 15–20 | Птушкi | 4–20 |
| Жабы | 5–14 | ||
| Млекакормячыя | |||
| Сурок | 6,5–9,0 | Каза | 3,0–5,5 |
| Суслiк | 5,5–8,5 | Малпа | 3,0–5,5 |
| Палёўка | 5,5–8,0 | Чалавек | 2,5–4,0 |
| Хамяк | 5,5–8,0 | Сабака | 2,5–4,0 |
| Кажан | 5,0–7,5 | Свiння | 2,5–3,5 |
Па меры ўскладнення арганiзацыi арганiзма радыеадчувальнасць яго ўзрастае. У межах аднаго арганiзма яна мяняецца ў залежнасцi ад знешнiх фактараў, унутранага стану, жыццёвага цыкла (у беспазваночных) i iнш. Для дарослых насякомых лятальнымi з'яўляюцца дозы 800–2000 Гр, эмбрыёнаў – 1–250 Гр, лiчынак ранняга ўзросту – 20–150, кукалак – 20–250 Гр. Генетычныя змяненнi ў элементах жывой прыроды адбываюцца пры дозах 0,1 Гр/год, прыметныя прызнакi прамянёвага паражэння ў найбольш адчувальных вiдаў жывёлы – 2 Гр/год, знiжэнне колькасцi жывёл назiраецца пры дозах, якiя складаюць 1–20% ЛД50/30. Экалагiчная мяжа дозы для экасiстэмы хваёвага лесу складае 30 Гр/год, лiставага – 300, большасцi травянiстых вiдаў – 400, прэснаводных – 300 Гр/год (Сушчэня Л. М., Пiкулiк М. Я., Пленiн Е. А.).
Радыенуклiды ў арганiзмы прадстаўнiкоў фауны паступаюць праз органы дыхання, страўнiкава-кiшэчны тракт i скурнае покрыва. Суадносiны iх паступлення звязаны з асаблiвасцямi арганiзмаў, канцэнтрацыяй i ўласцiвасцямi радыенуклiдаў. Шляхi мiграцыi радыенуклiдаў у бiясферы паказаны на рыс. 12.6.
Рыс. 12.6. Схема асноўных шляхоў мiграцыi радыенуклiдаў па элементах бiясферы
Уся разнастайнасць уздзеяння фауны на лясныя бiягеацэнозы зводзiцца да стварэння другаснай бiялагiчнай прадукцыi i бялковых кармоў, удзелу ў кругавароце асноўных бiягенных элементаў, механiчнай дзейнасцi. Хаця на долю бiямасы жывёлы ў агульнай структуры бiягеацэнозаў прыходзяцца сотыя долi працэнта, даследаванне заканамернасцей мiграцыi радыенуклiдаў у трафiчных ланцугах жывёлы з'яўляецца адным з важных i прыярытэтных кiрункаў у радыеэкалогii.
Непасрэднае апраменьванне i змяненне структуры ляснога бiягеацэнозу i мiкракл
Дата добавления: 2016-04-06; просмотров: 954;