Торможение — механизм формирования условных рефлексов. В
основе стадии специализации условного рефлекса лежит торможение. Торможение делят на два типа: 1) врожденное, безусловное, внешнее торможение; 2) приобретенное индивидуально, условное, внутреннее торможение. Внешнее торможение вызывается посторонним для данной условной реакции внешним стимулом и всегда начинается с ориентировочной реакции. Нейроны двигательной коры реагируют на внешние отвлекающие стимулы так же, как и на условные раздражения. Внешнее торможение вызывает безусловную ориентировочную реакцию и затормаживает условно-рефлекторную реакцию. Это осуществляется в результате включения в безусловную ориентировочную реакцию тех же нейронов коры, которые реагируют и на условный раздражитель.


 В зависимости от способа выработки условного рефлекса описывают разные виды внутреннего торможения: угасательное, диффе-ренцировочное, запаздывательное и условный тормоз. Угасательное торможение возникает после отмены подкрепления: условный рефлекс без подкрепления постепенно угасает. Дифференцировочное торможение вырабатывается на основе положительного условного рефлекса, когда в эксперимент вводится раздражение, близкое по параметрам к условному, но не сопровождаемое подкреплением. Запаздывательное торможение возникает при постепенном отставлении во времени безусловного подкрепления, скрытый период появления условного рефлекса увеличивается — рефлекс запаздывает. Условный тормоз вырабатывается при попеременном предъявлении подкрепляемого и неподкрепляемого условного раздражения, причем последнему предшествует добавочное раздражение. Именно добавочное раздражение приобретает способность тормозить следующий за ним условный раздражитель и становится условным тормозом.
В зависимости от способа выработки условного рефлекса описывают разные виды внутреннего торможения: угасательное, диффе-ренцировочное, запаздывательное и условный тормоз. Угасательное торможение возникает после отмены подкрепления: условный рефлекс без подкрепления постепенно угасает. Дифференцировочное торможение вырабатывается на основе положительного условного рефлекса, когда в эксперимент вводится раздражение, близкое по параметрам к условному, но не сопровождаемое подкреплением. Запаздывательное торможение возникает при постепенном отставлении во времени безусловного подкрепления, скрытый период появления условного рефлекса увеличивается — рефлекс запаздывает. Условный тормоз вырабатывается при попеременном предъявлении подкрепляемого и неподкрепляемого условного раздражения, причем последнему предшествует добавочное раздражение. Именно добавочное раздражение приобретает способность тормозить следующий за ним условный раздражитель и становится условным тормозом.
Все эти виды торможения выявляются с помощью экспериментальной процедуры, но основываются на реальных нейрофизиологических процессах. Например, исследование угасательного торможения на уровне одиночных нейронов мозга крыс при пищевом условном рефлексе показало, что самое быстрое угасание реакций при действии неподкрепляемых раздражителей наблюдается у нейронов ретикулярной формации. Реакции нейронов гиппокампа в тех же условиях угасают лишь частично, так же как и реакции нейронов гипоталамуса и преоптической области. Реакции корковых нейронов угасают медленно. Дифференцировочное торможение на уровне одиночных нейронов моторной коры развивается, в основном, в течение первых 5-7 проб, включающих дифференцировочный сигнал. Сначала нейроны реагируют одинаково как на условный, так и на дифференцировочный раздражители. При повторном тестировании появляются и нарастают различия в реакциях на эти раздражители и на конечном этапе выработки дифференцировки реакция на условное положительное раздражение превышает реакцию на дифференцировочное.
Все характерные для угашения и дифференцировки изменения в импульсной активности происходят в тех же структурах мозга, которые принимают участие в выработке условнорефлекторных реакций. Направление перестройки импульсной активности при внутреннем торможении противоположно тому, что наблюдается при выработке условного рефлекса. Внутреннее торможение с нейрофизиологической точки зрения — средство, блокирующее или изменяющее течение приобретенных индивидуальных поведенческих реакций.
Важным средством формирования условнорефлекторной деятельности является доминанта (А.А.Ухтомский), которая представляет собой временно господствующую рефлекторную систему, которая направляет работу нервных центров в данный момент. Если тот или иной центр мозга подготовлен к реакции, благодаря предварительным слабым влияниям внешних или внутренних раздражителей, т.е.
122


 возбудимость его повышена, то возбуждение нарастает в нем под влиянием импульсов, поступающих в центральную нервную систему, независимо от места и модальности раздражителя. Многократно усиленное в доминантном центре возбуждение тормозит все прочие рефлексы. Структурно-функционально доминанта представляет собой объединение нервных центров, состоящее из корковых, подкорковых, вегетативных и гуморальных систем. Доминанта — не топографически единый пункт возбуждения в центральной нервной системе, а синхронно работающее объединение центров с оптимальным для данной реакции стационарным возбуждением в разных этажах головного и спинного мозга и вегетативной нервной системе. Взаимодействие доминанты и условного рефлекса объясняет схема на рис.17.1.
возбудимость его повышена, то возбуждение нарастает в нем под влиянием импульсов, поступающих в центральную нервную систему, независимо от места и модальности раздражителя. Многократно усиленное в доминантном центре возбуждение тормозит все прочие рефлексы. Структурно-функционально доминанта представляет собой объединение нервных центров, состоящее из корковых, подкорковых, вегетативных и гуморальных систем. Доминанта — не топографически единый пункт возбуждения в центральной нервной системе, а синхронно работающее объединение центров с оптимальным для данной реакции стационарным возбуждением в разных этажах головного и спинного мозга и вегетативной нервной системе. Взаимодействие доминанты и условного рефлекса объясняет схема на рис.17.1.
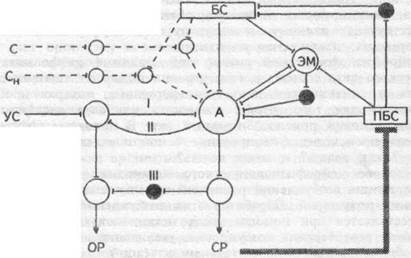
Рис.17.1. Схема взаимодействия доминанты (прерывистые линии) и условного рефлекса (сплошные линии). I — прямая условная связь, II — обратная связь — активирующая, III — тормозящая. Черные кружки — тормозные элементы. Тонкая пиния — влияние подкрепляющего стимула, заштрихованная — инструментальный условный рефлекс. С и Си — индифферентные стимулы; УС — условный стимул; БС — безусловный стимул, запускающий реакцию; ПБС — подкрепляющий безусловный стимул; А — афферентные дуги рефлексов; Э — эфферентные элементы рефлексов; ОР — ориентировочная реакция; СР — специфическая рефлекторная реакция, ЭМ — эмоция.
 123
123
 Функциональная архитектура условного рефлекса. Детальное изучение условнорефлекторной деятельности с использованием электрофизиологических данных в сопоставлении с вегетативными и двигательными компонентами условных реакций, привели к формулировке теории функциональной системы (П.К.Анохин) как основы понимания функциональной структуры условного рефлекса.
Функциональная архитектура условного рефлекса. Детальное изучение условнорефлекторной деятельности с использованием электрофизиологических данных в сопоставлении с вегетативными и двигательными компонентами условных реакций, привели к формулировке теории функциональной системы (П.К.Анохин) как основы понимания функциональной структуры условного рефлекса.
Поведение с позиции теории функциональных систем рассматривается как приспособительный акт любой степени сложности, в основе которого лежат следующие процессы: 1) афферентный синтез, 2) стадия принятия решения, 3) формирование акцептора результата действия, 4) формирование интеграла эфферентных возбуждений (эфферентный синтез), 5) получение полезного результата системы, 6) обратная афферентация о параметрах полученного реального результата, сопоставление их с ранее сформировавшимся акцептором результата действия.
Афферентный синтез — это первая стадия поведенческого акта, на которой решается вопрос: "Какой результат должен быть получен системой?". Афферентный синтез включает четыре основные компонента: доминирующая мотивация; обстановочная афферентация, соответствующая данному моменту; пусковая афферентация, а также информация, извлекаемая из памяти.
Нейрофизиологический смысл этой стадии формирования поведенческого акта состоит в том, что она позволяет тщательно обработать и синтезировать всю ту информацию, которая необходима организму для того, чтобы совершить наиболее адекватный для данных условий приспособительный акт. В процессе афферентного синтеза происходит формирование основ поведенческого акта: что делать? как делать? и когда делать?
В основе нейрофизиологического механизма этой стадии лежит конвергенция возбуждений различной модальности на нейронах коры больших полушарий. Обработка этих возбуждений нейронами коры осуществляется при помощи таких механизмов, как корково-подкорковая реверберация возбуждения, увеличение дискриминационной способности нейрона к частоте импульсаций, а также увеличение конвергентной емкости отдельного нейрона с обязательным участием ориентировочно- исследовательской реакции.
Стадия принятия решения является, по существу, стадией, на которой осуществляется формирование конкретной конечной цели, к которой стремится организм. Принятие решения является именно тем критическим пунктом, который переводит один системный процесс — афферентный синтез в другой системный процесс — в программу действий, и является тем переходным пунктом, после которого все комбинации возбуждений приобретают исполнительный характер. Проблема принятия решения на нейрональном уровне состоит из двух частей: 1) задача отдельного нейрона, 2) интеграция нейронов в единую систему.
Одним из наиболее интересных и сложных элементов функциональной системы является акцептор результата действия. Он обеспечивает прогнозирование признаков будущего результата и
124






 сравнение их при помощи обратной афферентации с параметрами реального результата системы.
сравнение их при помощи обратной афферентации с параметрами реального результата системы.
Представление об акцепторе основано на опытах, проводившихся по секреторно-двигательному методу условных реакций активного выбора со стороны подкрепления, на станке с двумя кормушками при подмене подкрепления. Этот прием "сюрпризной ситуации" с подменой подкрепления вызывал у животного бурную ориентировочную реакцию. Значит, в центральной нервной системе еще до получения животным реального результата, к которому оно стремится, формируется "образ" этого результата, а, следовательно, должен существовать и аппарат "подгонки" полученного результата и егопрообраза.
Функционирование акцептора результата действия изменяет устоявшиеся представления о работе центральной нервной системы по рефлекторному принципу. Формируясь на основе многостороннего механизма афферентного синтеза, акцептор результата действия отражает непоследовательное развитие явлений поведенческого акта. Он предвосхищает свойства того результата, который должен быть получен в соответствии с принятым решением, т.е. опережает ход событий в отношении между организмом и внутренней средой. Этот аппарат дает организму возможность исправить ошибку поведения.
На стадии эфферентного синтеза формируются центральные механизмы, которые обеспечивают получение определенного результата.
В ходе реализации целенаправленного поведения через звено обратной афферентации осуществляется постоянная оценка реально полученного результата с тем, который был запрограммирован в акцепторе результата действия. Результат этой оценки и определяет дальнейшее поведение организма. Если реально полученный результат соответствует прогнозируемому, то организм переходит к следующему этапу деятельности. При их несоответствии в аппарате сличения возникает рассогласование, активирующее ориентировочно-исследовательскую деятельность организма.
Функциональная система условного рефлекса имеет два принципиально важных свойства: 1) полезный приспособительный результат, 2) логическая операциональная архитектоника, представленная конкретными механизмами с обязательной обратной афферентацией в центральную нервную систему о результатах действия.
Память
Память — одно из основных свойств нервной системы, выражающееся в способности длительное время хранить информацию о событиях внешнего мира и реакциях организма, неоднократно выводить эту информацию в область сознания и поведения. Обучение и память — это две стороны одного процесса. При изучении обучения прежде всего исследуется механизм приобретения знаний, при изучении памяти — механизм хранения и использования этих знаний.




 Память человека и животных включает четыре характеристики — запоминание (усвоение) информации, ее сохранение, извлечение и воспроизведение. Запоминаются не только раздражители, исходящие из внешней или внутренней среды организма, но и вызванные ими ощущения и восприятия. Благодаря свойствам памяти, человек (и животные) организует во времени и пространстве функции, которые позволяют приобретать, сохранять и использовать индивидуальный опыт, причем, предыдущий опыт всегда влияет на последующее восприятие и запоминание.
Память человека и животных включает четыре характеристики — запоминание (усвоение) информации, ее сохранение, извлечение и воспроизведение. Запоминаются не только раздражители, исходящие из внешней или внутренней среды организма, но и вызванные ими ощущения и восприятия. Благодаря свойствам памяти, человек (и животные) организует во времени и пространстве функции, которые позволяют приобретать, сохранять и использовать индивидуальный опыт, причем, предыдущий опыт всегда влияет на последующее восприятие и запоминание.
Пространственная и временная организация памяти связана со многими структурами мозга. Прежде всего — это височная доля, гиппокамп и миндалина, а также связанные с ними структуры, мозжечок и кора больших полушарий, специфические и неспецифические таламические ядра. Роль этих структур была выявлена в экспериментах на животных и при различных травматических повреждениях у человека.
Медиальная височная область и гиппокамп участвуют в формировании и временном сохранении следов памяти, но не служат местами постоянного хранения информации. Больные с тяжелыми двусторонними поражениями гиппокампа не могли обучаться, не могли хранить в памяти то, что узнавали, не способны были вспомнить имя или лицо человека, которого только что видели. Память о событиях, происходивших до болезни или травмы, полностью сохранялась. Подтверждение роли гиппокампа в текущем запоминании было получено в опытах на крысах, которые после обучения переставали ориентироваться в лабиринте, если был поврежден гиппокамп. Если гиппокамп подвергнуть сильной электрической стимуляции, то у кролика не вырабатывается даже классический мигательный условный рефлекс.
Гиппокамп получает афферентацию от всех сенсорных систем, хотя и после множества переключений и, соответственно, после обработки информации в областях этих переключений. Миндалина и гипоталамус также связаны анатомически и функционально с сенсорными и висцеральными системами организма. Через гиппокамп, миндалину и гипоталамус проходят и частично переключаются пути центрифугальные, берущие начало от разных зон коры больших полушарий.
В опытах на обезьянах показано, что только одновременное удаление гиппокампа и миндалины уничтожает результаты недавнего обучения и лишает животное возможности дальнейшего обучения. Если удалялась только миндалина или только гиппокамп, то обучение новым формам поведения было возможно, хотя непосредственно предшествовавшее операции обучение практически было забыто животным.
Еще более существенна для организации памяти роль коры головного мозга. Медиальная часть височной коры связана с запоминанием текущих событий и наравне с гиппокампом обеспечивает сохранение у человека следов недавних событий. Люди с поврежденной височной долей (медиальная височная кора и гиппокамп)
126

 живут только в настоящем времени, у них нет недавнего прошлого. Существует в памяти только та часть жизни, которая прошла до операции или травмы. Все, что произошло в момент или после травмы, человек не помнит.
живут только в настоящем времени, у них нет недавнего прошлого. Существует в памяти только та часть жизни, которая прошла до операции или травмы. Все, что произошло в момент или после травмы, человек не помнит.
О роли различных областей коры мозга в процессах памяти косвенно свидетельствует то, что у животных, выращенных в условиях воздействия на них множества сенсорных раздражителей разной модальности и обучения (например, в лабиринте), слои коры толще, структура нейронов сложнее, чем у животных, выращенных в "обедненных" условиях. Найти определенное место или несколько мест в коре больших полушарий, где хранится та или иная информация или осуществляется только определенная функция памяти, пока не удалось. Делают заключение, что память широко распределена в различных областях мозга. И все-таки "преимущественные" функции структур мозга в организации памяти существуют, как это видно было для гиппокампа и височной коры мозга. Некоторая избирательность в отношении функций памяти характеризует таламические ядра и лобную кору.
У человека с повреждением дорсомедиального ядра таламуса в результате травмы память на события, предшествовавшие травме, полностью сохранялась, но возможность усвоения нового словесного материала практически исчезла, хотя запоминание лиц и определенных мест в окружающем пространстве частично сохранилось. Скорость забывания не словесной, а пространственной информации при повреждении дорсомедиального ядра таламуса была такая же, как у здоровых людей.
Поражения медиального таламуса в сочетании с дегенерацией нейронов лобной коры и мозжечка у человека приводят к неспособности решения задач в связи с невозможностью переучивания. Различные психологические приемы, используемые для улучшения запоминания у нормальных людей, не дают никакого результата при описанной выше патологии.
В сохранении результатов обучения двигательным условным рефлексам большое значение имеет мозжечок. Удаление зоны мозжечка, участвующей в моторном контроле двигательного мигательного рефлекса на звук, приводит к полному исчезновению у животных условного рефлекса. В то же время безусловный компонент рефлекса полностью сохраняется — вдувание воздуха в глаз вызывает мигание. Выработать условный рефлекс со стороны глаза, соответствующего оперированной стороне мозжечка, не удается. Это значит, что разрушение определенной области мозжечка ликвидировало не только следы обучения, но и блокировало образование новых следов. Разрушение глубинных мозжечковых ядер гибельно для следов памяти об обучении движениям. Изменения памяти, особенно ее эмоциональных аспектов, связаны с сохранностью миндалевидного комплекса (миндалины). Интактность лобных долей необходима для выполнения отсроченных реакций, сохранность височной коры является условием сохранения памяти как на относительно недавние, так и на отдаленные по времени события.
127

 У человека существует не менее трех различных типов памяти: "непосредственный отпечаток" сенсорной информации (иконинеская память), кратковременная и долговременная память. В зависимости от рецепторов, воспринимающих раздражения, выделяют зрительную, слуховую, осязательную, обонятельную, вкусовую и другие "памяти".
У человека существует не менее трех различных типов памяти: "непосредственный отпечаток" сенсорной информации (иконинеская память), кратковременная и долговременная память. В зависимости от рецепторов, воспринимающих раздражения, выделяют зрительную, слуховую, осязательную, обонятельную, вкусовую и другие "памяти".
Иконическая память удерживает точную и полную картину, воспринимаемую органами чувств, то есть образ предмета. Длительность хранения образа 0,1-0,5 с. Емкость ее ограничена 3-5 элементами. Этот тип памяти связывают с чисто сенсорными процессами — последействием в периферических и центральных звеньях, связанным с иннерционностью периферического процесса, например, разложением зрительного пигмента.
Кратковременная память удерживает не точную копию предмета, события, явления, а их частичное отображение, емкость ее невелика — 7±2 предъявляемых элемента. Длительность сохранения следов от 5 до 60 с. Запоминание связано с повторением, например, 5-7 слов или цифр, что позволяет сохранять информацию более длительное время.
Долговременная память удерживает огромный объем информации. Все, что содержится в памяти более одной минуты, переводится в систему долговременной памяти, где и сохраняется часами, а иногда на протяжении всей жизни. Основой функционирования системы, которая имеет дело с большим количеством запечатленной в памяти информации, является не физическая емкость, а способность отыскать ответ на поставленный перед такой системой вопрос. Именно поэтому, долговременная память составляет основное звено в организации целенаправленного поведения, обеспечивая хранение, извлечение и воспроизведение информации из внешней и внутренней среды организма.
Переход от кратковременной к долговременной памяти — это преобразование процесса получения информации в процесс ее сохранения. Одна из систем мозга, обеспечивающих подобное преобразование — гиппокамп. В процессах перехода информации от кратковременного к длительному хранению участвует внимание (сознательный компонент), которое контролируется ретикулярной формацией мозга. Часть данных запоминается и в отсутствие внимания — это непроизвольное запоминание (подсознательный компонент). Запоминание или консолидация следов памяти осуществляется с участием медиальной височной доли и гиппокампа. После консолидации следов данные становятся постоянным содержанием долговременнрй памяти.
Запоминание осуществляется двумя способами — процедурным и декларативным. Процедурное запоминание связано с получением и хранением знаний о том, как надо действовать, а декларативное — о том, что составляет основу действия. Классические условные рефлексы — это способы- приобретения и закрепления знаний о том, как надо действовать. Процедурная память связана только с теми нервными структурами, которые непосредственно участвуют в усвоенных действиях. Познавательные процессы, связанные с осознанием действий, в простейшем случае — это произвольные осознанные
128


 тью которой является речь и письмо. Помимо первичных, существуют также вторичные сенсорные и моторные поля, расположенные в непосредственной близости к первичным зонам. Взаимодействие перечисленных корковых зон осуществляется как за счет транскортикальных ассоциативных связей, так и корково-талами-ческих и соответствующих таламо-корковых связей.
тью которой является речь и письмо. Помимо первичных, существуют также вторичные сенсорные и моторные поля, расположенные в непосредственной близости к первичным зонам. Взаимодействие перечисленных корковых зон осуществляется как за счет транскортикальных ассоциативных связей, так и корково-талами-ческих и соответствующих таламо-корковых связей.
Распределение в правом и левом полушарии мозга такой высокоспециализированной функции человека как речь, глубоко асимметрично. Лингвистические способности человека определяются, преимущественно, левым полушарием. Три взаимосвязанные речевые зоны, расположенные в задней височной области, нижней центральной извилине и в дополнительной моторной коре левого полушария, действуют как единый речевой механизм. Пути осуществления кооперации различных областей и их функции представлены на рис.17.3.
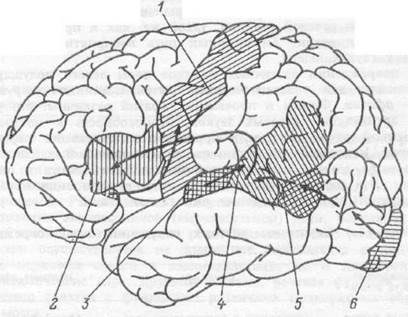
Рис. 17.3. Корковые пути, связывающие различные слухо- речевые зоны.
1 — моторная кора, 2 — зона Брока, 3 — первичная слуховая кора, 4 — зона Вернике, 5 — угловая извилина, 6 — первичная зрительная кора.
 После того, как акустическая информация, заключенная в слове, обрабатывается в слуховой системе и в других "неслуховых" образованиях мозга, она поступает в первичную слуховую кору. Однако, для понимания человеком смысла речи и выработки программы
После того, как акустическая информация, заключенная в слове, обрабатывается в слуховой системе и в других "неслуховых" образованиях мозга, она поступает в первичную слуховую кору. Однако, для понимания человеком смысла речи и выработки программы
141
 речевого ответа необходима дальнейшая обработка полученной информации. Она осуществляется в зоне Вернике, расположенной в височной области, в непосредственной близости к первичной слуховой коре. Именно здесь обеспечивается понимание смысла поступившего сигнала — слова. Для произнесения слова необходимо, чтобы активировалось его представительство в зоне Брока, расположенной в третьей лобной извилине. Активация зоны Брока после понимания смысла речи, благодаря участию зоны Вернике, обеспечивается группой волокон, называемой дугообразным пучком. В зоне Брока сведения, поступившие из зоны Вернике, приводят к возникновению детальной программы артикуляции. Реализация этой программы осуществляется через активацию лицевой проекции моторной коры, управляющей речевой мускулатурой и связанной с зоной Брока короткими волокнами. Если воспринимается письменная речь, то сначала включается первичная зрительная кора. После этого информация о прочитанном слове поступает в угловую извилину, которая связывает зрительную форму данного слова с его акустическим аналогом в зоне Вернике. Дальнейший путь, приводящий к возникновению речевой реакции, такой же, как и при чисто акустическом восприятии. Аналогичный путь восприятия письменной речи и у глухих людей.
речевого ответа необходима дальнейшая обработка полученной информации. Она осуществляется в зоне Вернике, расположенной в височной области, в непосредственной близости к первичной слуховой коре. Именно здесь обеспечивается понимание смысла поступившего сигнала — слова. Для произнесения слова необходимо, чтобы активировалось его представительство в зоне Брока, расположенной в третьей лобной извилине. Активация зоны Брока после понимания смысла речи, благодаря участию зоны Вернике, обеспечивается группой волокон, называемой дугообразным пучком. В зоне Брока сведения, поступившие из зоны Вернике, приводят к возникновению детальной программы артикуляции. Реализация этой программы осуществляется через активацию лицевой проекции моторной коры, управляющей речевой мускулатурой и связанной с зоной Брока короткими волокнами. Если воспринимается письменная речь, то сначала включается первичная зрительная кора. После этого информация о прочитанном слове поступает в угловую извилину, которая связывает зрительную форму данного слова с его акустическим аналогом в зоне Вернике. Дальнейший путь, приводящий к возникновению речевой реакции, такой же, как и при чисто акустическом восприятии. Аналогичный путь восприятия письменной речи и у глухих людей.
При повреждении различных участков коры левого полушария и соединяющих эти участки нервных путей возникают нарушения речи — афазии. Формы и проявления афазий различны: это нарушение артикуляции речевых звуков, неспособность к построению осмысленной речи, даже если произнесение звуков не нарушено, это также неспособность понимать устную речь. В таблице 17.4суммированы данные о роли различных областей мозга в речевой функции. Эта таблица базируется на результатах оценки патологии речи (афазий) при повреждении разных зон мозга.
Таблица 17.4Симптомы речевых нарушений при повреждении различных зон мозга
 Поврежденная область мозга
Поврежденная область мозга
Симптомы
Название афазии
 Лобная доля левого полушария, третья лобн!Я извилина — зона Брока.
Лобная доля левого полушария, третья лобн!Я извилина — зона Брока.
Височная доля левого полушария, задняя часть первой височной извилины или зона Вернике.
Затруднения в произнесении слов. Понимание речи, чтение и письмо не нарушены. Больной осознает свой дефект.
Затруднения в произнесении звуков и слов отсутствуют, речь беглая, но бессмысленная, содержит несуществующие слова, ритм, интонации, грамматические формы сохранены. Понимание речи, чтение и письмо сильно нарушены. Больной не осознает дефект.
Афазия Брока
Афазия Вернике
 142
142

 Таблица 17.4 (продолжение)
Таблица 17.4 (продолжение)


 Поврежденная область мозга
Поврежденная область мозга
 Нервные волокна, соединяющие третью лобную и заднюю часть височной извилины (зоны Брока и Вернике)
Нервные волокна, соединяющие третью лобную и заднюю часть височной извилины (зоны Брока и Вернике)
Область между первичной слуховой корой и зоной Вернике
Угловая извилина — место соединения височной, теменной и затылочной долей левого полушария.
Обширное поражение левого полушария.
Симптомы
Речь беглая, бессмысленная, повторение фразы затруднено. Сохранена некоторая способность к пониманию речи и чтению.
Понимание устной речи нарушено. Устная речь и письмо не нарушены. Понимание письменной речи сохранено.
Неспособность назвать предметы, имя человека, нарушение памяти на слова в их связи с предметами.
Нарушение всех языковых функций.
Название афазии
Проводниковая афазия
Словесная глухота
Анатомическая афазия (аномия — неспособность вспомнить слово и дать название предметам и явлениям).
Глобальная афазия



 Корковые отделы левого полушария играют специфическую роль в восприятии, запоминании и воспроизведении речевого материала. Именно эти зоны необходимы для полноценного осуществления речевой функции как единого сенсорного, мыслительного и моторного процесса. Расположенные спереди зоны особенно важны для осуществления экспрессивной (выразительной) речи, расположенные сзади — для восприятия смысла речи. Взаимодействие речевых корковых зон осуществляется не только по горизонтали с помощью корково- корковых связей и взаимодействий, но и по вертикали, через таламические ядра. Другими словами, речевая функция непосредственно связана с функциями различных подкорковых образований мозга.
Корковые отделы левого полушария играют специфическую роль в восприятии, запоминании и воспроизведении речевого материала. Именно эти зоны необходимы для полноценного осуществления речевой функции как единого сенсорного, мыслительного и моторного процесса. Расположенные спереди зоны особенно важны для осуществления экспрессивной (выразительной) речи, расположенные сзади — для восприятия смысла речи. Взаимодействие речевых корковых зон осуществляется не только по горизонтали с помощью корково- корковых связей и взаимодействий, но и по вертикали, через таламические ядра. Другими словами, речевая функция непосредственно связана с функциями различных подкорковых образований мозга.
Итак, функциональная асимметрия мозга в связи с механизмами речи проявляется следующим образом. Тональный слух идентичен для обоих полушарий. Участие левого полушария необходимо для обнаружения и опознания артикулированных звуков речи, а правого — для опознания интонаций, транспортных и бытовых шумов, музыкальных мелодий. Восприятие и генерация звуков речи, а также более высокий уровень общей речевой активности, обеспечиваются левым полушарием, а улучшение выделения сигнала из шума — правым. Правое полушарие не способно реализовывать команду для продуцирования речи, но оно обеспечивает понимание устной речи и написанных слов. Понимание речи, осуществляемое правым полушарием, ограничено конкретными именами существительными, в
143
 меньшей степени — отглагольными существительными, еще в меньшей степени — глаголами. Правое полушарие обеспечивает понимание эмоционального содержания интонаций, опознание по голосу, участвует в модуляции частот голоса.
меньшей степени — отглагольными существительными, еще в меньшей степени — глаголами. Правое полушарие обеспечивает понимание эмоционального содержания интонаций, опознание по голосу, участвует в модуляции частот голоса.
Контроль результата деятельности речевой системы.Для оценки успешного выполнения той или иной моторной поведенческой программы, в том числе и речи, необходим контроль ее реализации как в процессе выполнения, так и по конечному результату. Такая оценка осуществляется мозгом человека, благодаря системам с обратными связями. У человека существуют три сенсорных канала получения информации об успешной реализации речевого процесса: (1) слуховой, (2) проприоцептивный и кинестетический, (3) зрительный.
Точность воспроизведения речи, то есть соответствия акустической формы речевого сигнала его акустическому образу, сформированному в процессе обучения, контролирует слуховая обратная связь. Она начинается в слуховой височной зоне и проходит в центробежном направлении параллельно путям и центрам афферентной слуховой системы вплоть до волосковых клеток улитки внутреннего уха. Точность воспроизведения речи контролируется также оценкой аффе-рентации от проприоцептивных и кинестетических рецепторов, расположенных в мышцах и суставах речеобразующих органов. Полученная от рецепторов соматосенсорной системы афферентация сравнивается с хранящимся в памяти образом правильного двигательного акта и оценивается по степени соответствия эталона его реализации в движении. В механизмах проприоцептивного контроля участвуют кора третьей лобной извилины левого полушария и нижняя моторная кора обоих полушарий. Контроль конечного результата влияния экспрессивной речи на слушателя реализуется по зрительному и слуховому каналам афферентации. Зрительное восприятие обеспечивает человеку оценку реакции слушателя на произнесенные звуки. Это позволяет определить, достигло ли речевое сообщение той цели, которую имел в виду говорящий. Слуховой канал позволяет говорящему дать оценку результата воздействия речи по речевому ответу слушателя. Эти виды контроля требуют участия многих структур мозга.
Раздельный контроль исполнения (качества речевой продукции) и конечного результата (смыслового содержания высказывания) дает возможность человеку выявить причины неудачи речи как способа коммуникации. Двойной контроль речевого процесса с помощью обратных связей позволяет отделить (1) плохое осуществление двигательной программы, лежащей в основе реализации речи как акустического явления, от (2) полной неадекватности речевого акта стоящей перед ним" цели, например, невозможности изменить поведение партнера или получить ответ на вопрос.
Говорящий и слушающий в процессе генерации и восприятия звуковой речи составляют единую систему. Коммуниканты, как части этой системы, осуществляют в процессе общения ряд общих функций. К их числу относятся, например, анализ акустических
144


 свойств речи, выделение речевого сообщения из шума, понимание смысла высказывания и его эмоционально-аффективного содержания. В диалоге, во всяком случае на определенных ограниченных временных отрезках слухоречевого взаимодействия, одна сторона (говорящий) выполняет одновременно две, а другая (слушающий) — одну функцию. Говорящий реализует семантическую программу речи в определенной грамматической и акустической форме. Параллельно он осуществляет двойной контроль качества речевой продукции: говорящий использует для контроля результата акустическую обратную связь, проприоцептивный и зрительный контроль параллельно. Для слушателя главной задачей является понимание смысла высказывания.
свойств речи, выделение речевого сообщения из шума, понимание смысла высказывания и его эмоционально-аффективного содержания. В диалоге, во всяком случае на определенных ограниченных временных отрезках слухоречевого взаимодействия, одна сторона (говорящий) выполняет одновременно две, а другая (слушающий) — одну функцию. Говорящий реализует семантическую программу речи в определенной грамматической и акустической форме. Параллельно он осуществляет двойной контроль качества речевой продукции: говорящий использует для контроля результата акустическую обратную связь, проприоцептивный и зрительный контроль параллельно. Для слушателя главной задачей является понимание смысла высказывания.
Одной из широко распространенных разновидностей речевой коммуникации является язык жестов. Это язык, использующий зрительно-пространственные возможности человеческого мозга. Это формальный язык со сложным словарем и грамматической структурой. Каждый знак представляет собой "букву", "слог" или "слово" — в зависимости от типа языка. Клинические наблюдения свидетельствуют о том, что у нормально слышащих и говорящих людей, пользующихся языком жестов, повреждение левого полушария в областях, связанных со звуковой речью, приводит к неспособности активно пользоваться языком жестов и понимать его. У тех глухонемых, которые начали пользоваться языком жестов в раннем детском возрасте, левое полушарие доминирует и при решении зрительно-пространственных задач. Значит, языковые функции реализуются сходными механизмами независимо от того, в какой форме речь используется — звуковой, письменной или жестовой.
Мышление— процесс познавательной деятельности человека, характеризующийся обобщенным и опосредованным отражением внешнего мира и внутренних переживаний. Первый этап в организации мышления у детей состоит в построении сенсомоторных схем (до 2-х лет). Сенсомоторная схема — это выполнение организованной последовательности действий, составляющих определенную форму поведения (ходьба, еда, речь и пр.). Сенсомоторная схема соотносит сенсорную информацию с моторными (мышечными) действиями. В формировании сенсо- моторных схем ведущая роль принадлежит таламо-кортикальным системам головного мозга. С развитием речи и появлением способности мысленно активировать сенсорно- моторные схемы, не совершая действия, формируется первая фаза человеческого мышления (2-7 лет).
Основной особенностью первой фазы человеческого мышления становится способность ребенка предсказывать результат действия, не производя его фактически. В этот период ребенок уже хорошо знает, что случится, если, например, бросить чашку со стола на пол, или дернуть кошку за хвост, или прыгать по лужам и т.д. Действие, однако, остается основным элементом мышления ребенка в этом возрасте. Если попросить ребенка дать определение какому-либо бытовому предмету или понятию, то его ответ будет содержать дей-
145




 ствие: стул — это то, на чем сидят, стол — это то, на чем едят, прогулка — это то, где бегают и т.д. В этот период развивается речь, которая первоначально базируется также на сенсорно-моторной схеме: слушаю — повторяю. При развитии речи сенсо-моторная схема получает название — слово. В период 2-7 лет бурное развитие претерпевает височная и моторная области коры головного мозга.
ствие: стул — это то, на чем сидят, стол — это то, на чем едят, прогулка — это то, где бегают и т.д. В этот период развивается речь, которая первоначально базируется также на сенсорно-моторной схеме: слушаю — повторяю. При развитии речи сенсо-моторная схема получает название — слово. В период 2-7 лет бурное развитие претерпевает височная и моторная области коры головного мозга.
Вторая фаза — способность к логическому рассуждению и использование, конкретных понятий в пределах реальных событий. В этот период (7-10 лет) активируются корково- корковые ассоциативные связи. Третья фаза — способность к формальным операциям, к абстракциям, к оценке гипотез (11-15 лет). Считают, что в этот период завершается формирование связей лобной коры с другими отделами мозга.
Мысленное моделирование человеком различных событий составляет сущность его мышления. Человек оценивает свои действия, ведущие к поставленной им цели, условия, которые приводят к успешному результату. Причем, последовательность событий может моделироваться в любом направлении, мысленные действия могут совершаться в разных точках выбора решения. Например, человек может начинать рассмотрение цепи событий и действий с желаемого результата и двигаться назад — в направлении начальных действий, мысленно выявлять, какие из них ведут к цели, находить условия, которые следует соблюсти для ее достижения.
Мышление имеет, как минимум, два аспекта: распознавание (принятие решения) и устойчивое сохранение поиска (стратегия решения задач). Мышление, как процесс принятия решения, требует участия височных и лобных отделов коры больших полушарий. Мышление, как поиск, осуществляется при участии, преимущественно, задних (теменно-затылочных) отделов коры мозга, а соответствие решения выработанному критерию (стратегия) реализуется при участии фронтальных, височных и лимбических отделов мозга. Структурные предпосылки мышления считают связаными с корой больших полушарий, преимущественно, с теми ее областями, которые объединены общим названием ассоциативная кора. В отличие от специфических, первичных корковых проекций различных сенсорных систем, ассоциативная кора является местом интеграции информации, поступающей из первичных проекций. Кроме того, считают, что в ассоциативных областях текущие сенсорные данные объединяются с информацией, содержащейся в памяти. Например, ассоциативные поля теменной зоны объединяют сведения, приходящие от первичных корковых проекций кожи, мышц, сухожилий, суставов, со слуховой и зрительной информацией, поступающей от височной и затылочной коры. Интеграция всех сенсорных данных со следами памяти (участие височной коры и гиппокампа) дает возможность человеку оценивать положение тела и головы в пространстве. Включение в оценку пространственных отношений лобных отделов коры позволяет человеку интерпретировать сенсорные раздражения в зависимости от конкретной ситуации. Именно ассоциативные поля коры лобной доли мозга имеют особое значение в интерпретации раздражителей и событий. Благодаря двусторонним
146

 связям лобной коры и лимбической системы, в систему оценки ситуации включаются эмоции. Кроме того, лобная кора ответственна за выбор целей и прогнозирование событий.
связям лобной коры и лимбической системы, в систему оценки ситуации включаются эмоции. Кроме того, лобная кора ответственна за выбор целей и прогнозирование событий.
Решающая роль лобных долей мозга в планировании и решении задач подтверждается сведениями о поведении людей с повреждениями этих областей коры. Они оказывались неспособными к решению задач в изменившейся ситуации или к осуществлению последовательных действий. То есть, одна и та же задача не могла быть решена, если ставилось дополнительное условие. Кроме того, использование языка, совершенно необходимое для многих мыслительных действий, возможно только при совместной работе лобных и височных долей (зоны Брока и зоны Вернике).
Билатеральная (полушарная) организация мозга — это еще один анатомический аспект, необходимый для целостного понимания того, как организовано мышление. Проникновение в процессы, связанные с мышлением, обусловлено возможностью клинического и физиологического исследования людей с повреждениями мозга, в особенности тех, у кого полушария оказались разделенными друг от друга — расщепленный мозг.
Человек имеет два полушария — правое и левое, выполняющие разные функции, но совместно обеспечивающие целенаправленное поведение. Полушария связаны между собой пучками волокон, самым мощным из которых является мозолистое тело. Правое полушарие контролирует и регулирует сенсомоторные и двигательные функции левой половины тела, левое — правой. То, что многие функции левого и правого полушарий головного мозга различны, было обнаружено у людей с расщепленным мозгом после травм или хирургических операций, выполненных в лечебных целях при тяжелых формах эпилепсии.
Каждое полушарие обладает собственными ощущениями, восприятием, мыслями и идеями, характеризуется разной эмоциональной оценкой идентичных событий. Каждое полушарие располагает собственной цепью воспоминаний и усвоенных знаний, недоступных для другого полушария. В определенных отношениях каждое полушарие имеет отдельное, собственное мышление. Левое — речевое, правое — зрительно-пространственное. Левое полушарие обрабатывает информацию аналитически и последовательно, правое — одновременно и целостно. Каждое полушарие вносит свой уникальный вклад в мышление и сознание (рис. 17.4). Ниже приводятся два примера различных функций правого и левого полушарий.
Женщина, у которой было перерезано мозолистое тело по поводу тяжелой эпилепсии, для предотвращения генерализации припадков, проходила зрительное тестирование с помощью тахистоскопа — прибора, позволяющего предъявлять на экране зрительное изображение в течение точно контролируемого, обычно короткого, времени (десятые доли секунды). В центре экрана находится черная точка, на которую должна смотреть испытуемая, не отрывая взгляда, так называемая, точка фиксации взора. На десятую долю секунды справа от точки появляется изображение предмета. За столь короткое время
147



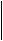





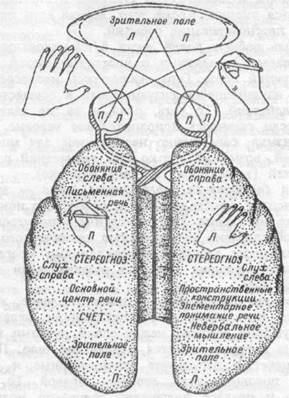
Рис.17.4. Основные функции правого и левого полушарий мозга человека.
 испытуемая не успевает перевести взор от точки на изображение. Смысл подобной стимуляции состоит в том, чтобы изображение попало в одно полушарие мозга, в данном случае — в левое. Если на экране справа появляется ложка, то оперированная испытуемая отвечает, что она видела "ложку". Если изображение ложки появляется слева от точки фиксации взора, то есть попадает в правое неречевое полушарие, то испытуемая отвечает: "Не видела ничего". Однако, если ей предлагалось выбрать левой рукой наощупь среди нескольких предметов тот, который возможно мелькнул на экране, но не был ею опознан, то она выбирала ложку. То есть, правое полушарие было способно опознавать предмет, но не смогло его назвать, т.к. связи с речевым левым полушарием были рассечены. На вопрос, что она держит в руках, оперированная отвечала: "карандаш". Больная "знает", и наощупь, т.е. пространственным способом, опознает, что она видела, но не может правильно определить предмет словесно.
испытуемая не успевает перевести взор от точки на изображение. Смысл подобной стимуляции состоит в том, чтобы изображение попало в одно полушарие мозга, в данном случае — в левое. Если на экране справа появляется ложка, то оперированная испытуемая отвечает, что она видела "ложку". Если изображение ложки появляется слева от точки фиксации взора, то есть попадает в правое неречевое полушарие, то испытуемая отвечает: "Не видела ничего". Однако, если ей предлагалось выбрать левой рукой наощупь среди нескольких предметов тот, который возможно мелькнул на экране, но не был ею опознан, то она выбирала ложку. То есть, правое полушарие было способно опознавать предмет, но не смогло его назвать, т.к. связи с речевым левым полушарием были рассечены. На вопрос, что она держит в руках, оперированная отвечала: "карандаш". Больная "знает", и наощупь, т.е. пространственным способом, опознает, что она видела, но не может правильно определить предмет словесно.
В другом исследовании с применением тахистоскопа испытуемой с расщепленным мозгом предъявляли четыре фотографии разных
148

 людей и называли их имена. Затем испытуемая садилась перед экраном, фиксировала взор на точку в центре экрана. Справа от точки на экране появляется половина фотографии лица одного человека, слева — половина изображения лица другого. Испытуемая называет имя человека, фотография половины лица которого попала в правое поле зрения и, соответственно, в левое полушарие. То есть, на вопрос отвечает левое, речевое полушарие. Потом части фотографии лица человека предъявляются испытуемой еще раз. Но на этот раз ее просят показать, а не назвать человека, лицо которого она видела. Испытуемая показывает на фотографии лицо, половина изображения которого попала в левое поле зрения, т.е. в правое, не говорящее полушарие, но способное оценить зрительно-пространственные соотношения.
людей и называли их имена. Затем испытуемая садилась перед экраном, фиксировала взор на точку в центре экрана. Справа от точки на экране появляется половина фотографии лица одного человека, слева — половина изображения лица другого. Испытуемая называет имя человека, фотография половины лица которого попала в правое поле зрения и, соответственно, в левое полушарие. То есть, на вопрос отвечает левое, речевое полушарие. Потом части фотографии лица человека предъявляются испытуемой еще раз. Но на этот раз ее просят показать, а не назвать человека, лицо которого она видела. Испытуемая показывает на фотографии лицо, половина изображения которого попала в левое поле зрения, т.е. в правое, не говорящее полушарие, но способное оценить зрительно-пространственные соотношения.
Итак, исследования лиц с расщепленным мозгом свидетельствуют о том, что левое полушарие отвечает за язык и речь, а правое управляет пониманием и навыками, связанными с пространственным и зрительным восприятием. В то же время, правое полушарие обладает способностью понимать речь, но не может ее программировать. Таким образом, если схематизировать функции полушарий в отношении мышления, то оказывается, что левое и правое полушария в равной степени способны к распознаванию стимулов внешнего мира, но пользуются разными способами или стратегиями решения задачи и имеют разные возможности в выражении результатов решения — языковую для левого полушария и пространственно-зрительную для правого полушария.
Между полушариями головного мозга существуют не только функциональные, но и структурные различия, особенно в височной области. В частности, участок коры височной доли, примыкающий и перекрещивающийся с зоной Вернике, у подавляющего большинства людей (70 из 100 исследованных посмертно) слева значительно больше, чем справа.
Структурная ассиметрия характерна не только для мозга взрослого человека, владеющего речью, но и для мозга человеческого плода. Асимметрия правой и левой половины мозга зарегистрирована с помощью вызванных потенциалов также у новорожденных. Асимметрия выявилась и при изучении ископаемых черепов неандертальского человека. Видимо, асимметрия полушарий составляет часть генетической программы человека.
При удалении височных зон левого полушария у взрослого больного человека развивается необратимый дефект речи — афазия. Однако, подобная операция у детей младенческого возраста не приводит к нарушениям речи. Более того, специальные психологические тесты не выявляют различий в развитии интеллекта между детьми, перенесшими операцию, и их нормальными сверстниками. Нет различий также и в развитии детей с право- и левосторонней гемисферэктомией (удаление большей части полушария). Причиной компенсации функций при повреждении мозга является его пластичности.
Два полушария мозга обладают специализированными функциями, но в интактном мозге они взаимодействуют и обуславливают высо-
149


 кую приспособляемость человека к окружающим условиям среды, огромную пластичность его поведения, обеспечивая целостное восприятие внешнего мира и самого себя.
кую приспособляемость человека к окружающим условиям среды, огромную пластичность его поведения, обеспечивая целостное восприятие внешнего мира и самого себя.
Дата добавления: 2016-03-27; просмотров: 1271;