водно-солевого обмена и гомеостатическая функция почек
Водно-солевой гомеостазис является одной из важнейших характеристик внутренней среды организма. От состояния водно-солевого обмена зависят фундаментальные свойства клеток и тканей (энергетический и пластический метаболизм, возбудимость, проводимость
и сократимость, секреторная способность и т.п.) и, соответственно, функции практически всех физиологических систем. Основные характеристики водно-солевого обмена — количество поступающих в организм жидкости и солей, объемы внеклеточных и внутриклеточных водных пространств, содержание в них отдельных электролитов и осмотическое давление, наконец, выделение воды и солей почками — имеют взаимосвязанные комплексные механизмы регуляции. Эти механизмы легко проследить на примерах изменения приема жидкости или ее потерь, сдвигов баланса натрия и калия, изменения объема крови.
Ограничение приема воды (рис. 12.4) приводит к дегидратации внутренней среды организма, при этом имеет место повышение осмотического давления плазмы крови и внеклеточной жидкости, что
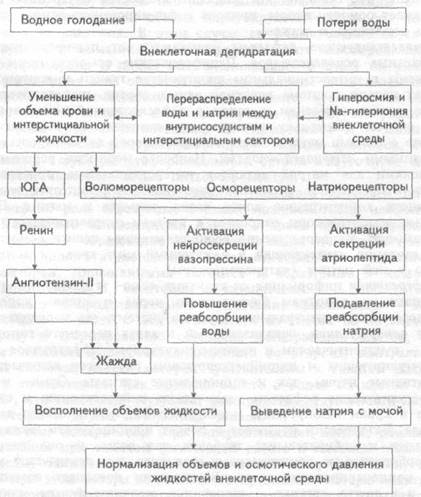
Рис. 12.4. Гомеостатическая регуляция водно-солевого обмена при дефиците воды и дегидратации.
523
дало основание именовать этот вид дегидратации гипертонической или истинной. Повышение осмотического давления плазмы крови вызывает перераспределение жидкости и электролитов между кровью, внеклеточной жидкостью и лимфой, а также между внеклеточной и внутриклеточной средой направленное на нормализацию сдвига осмотического давления. Начальные механизмы регуляции заключаются в буферировании белками и эритроцитами катионов; образовании с ними сложных химических соединений, с измененными осмотическими свойствами и гидрофильностью; диффузии осмотически активных веществ из крови в тканевую жидкость и лимфу; осмотическом токе воды в противоположном направлении; усилении всасывания воды в кишечнике и т.п. При небольших физиологических уровнях дегидратации этих механизмов бывает достаточно для обеспечения водно-солевого гомеостазиса.
Повышенное осмотическое давление жидкостей внутренней среды раздражает осморецепторы, приводя к формированию осморегулиру-ющих рефлексов и жажды.
Осморегулирующие рефлексы начинаются от периферических и центральных осморецепторов. Периферические осморецепторы расположены в интерстициальном пространстве тканей и кровеносных сосудах печени (система воротной вены), сердца, пищеварительного тракта, почек, селезенки, а также в рефлексогенной зоне каротид-ного синуса. Поскольку осмотическое давление преимущественно связано с ионами натрия, часть осморецепторов является специализированными натриорецепторами. Наиболее мощными рецептивными полями для натрия являются интерстициальные пространства печени и предсердий. Описаны также тканевые рецепторы, воспринимающие концентрацию ионов калия, кальция и магния. Центральные осморецепторы относятся к клеткам супраоптического ядра гипоталамуса, непосредственно воспринимающим сдвиги осмотического давления притекающей в межуточный мозг крови и концентрацию в ней натрия (натриорецепторы).
Афферентная информация от тканевых осмо- и натриорецепторов поступает по волокнам блуждающего нерва и задних корешков спинного мозга в центральную нервную систему, где направляется в центр осморегуляции, локализованный в ядрах переднего гипоталамуса — супраоптическом и паравентрикулярном. Эфферентное звено осморегулирующих и натрийрегулирующих рефлексов включает как вегетативные нервы, так и гормональные сигналы. Осмо- и на-трийрегулирующие рефлексы увеличивают нейросекрецию и содержание в крови вазопрессина и секрецию натриуретического атрио-пептида. Вазопрессин повышает в почках проницаемость дистальных канальцев и собирательных трубочек для воды, и она пассивно реабсорбируется по осмотическому градиенту, т.е. реализуется механизм концентрирования мочи. Вазопрессин повышает всасывание воды и в других органах — кишечнике, желчном пузыре, слюнных и пищеварительных железах. Натриуретический атриопептид, как и в меньшей степени вазопрессин, ведут к повышенному выведению натрия из крови, что устраняет увеличение осмотического давления.
524
Важнейшей приспособительной реакцией организма при дегидратации является жажда. Жажда — это субъективное ощущение, возникающее при абсолютном или относительном (к содержанию натрия) дефиците воды и приводящее к поведенческой реакции приема воды. Таким образом, жажда является одной из основных биологических мотиваций (т.е. побуждений к деятельности), обеспечивающих поддержание жизнедеятельности. Формирование чувства жажды связано с возбуждением питьевого центра, локализованного в структурах гипоталамуса и лимбики. Возбуждение центра обусловлено несколькими причинами.
Во-первых, при повышении осмотического давления крови и внеклеточной жидкости, уровня натрия в них, происходит раздражение центральных и периферических осморецепторов и рефлекторное возбуждение центра.
Во-вторых, возбуждение центра и возникновение жажды имеет место и при уменьшении объема циркулирующей крови без сдвига осмотического давления. В этом случае ее формирование обусловлено двумя механизмами: 1) волюморефлексом от волюморецепторов правого предсердия и полых вен, возникающим в ответ на уменьшение растяжения этих областей низкого давления сниженным объемом крови; 2) уменьшенный объем крови ведет к активации юкс-тагломерулярного аппарата почек и секреции ренина, что увеличивает в крови уровень ангиотензина-II. Ангиотензин-II — один из мощных дипсогенных (от греческого dipsa — жажда) факторов, стимулирующий структуры субфорникального органа межуточного мозга, который является частью питьевого центра.
В-третьих, в механизме возбуждения питьевого центра и генезе жажды играют роль образуемые в самой мозговой ткани нейропеп-тиды. Повышение концентрации натрия в крови при дегидратации влечет за собой образование в мозговой ткани натриуретического пептида, подобного атриопептиду, также относящегося к дипсоген-ным факторам. Дипсогенными являются и образуемые в мозговой ткани ангиотензин-II, окситоцин и вазопрессин.
Выраженное и продолжительное ограничение приема воды — водное голодание — ведет к выходу жидкости из клеток во внеклеточную среду и формированию состояния дегидратации клеток, что вызывает тяжелые расстройства, особенно со стороны центральной нервной системы, клетки которой чувствительны к сдвигам водно-солевого гомеостазиса. Как правило вместе с водой из клеток выходит К+, что еще более усугубляет расстройства деятельности нервной системы и других возбудимых тканей.
Избыточные потери воды (при гипервентиляции легких, обильном потоотделении при тяжелой физической работе и высокой температуре среды) также ведут к гипертонической гипергидратации, поскольку при этом потери воды намного превышают потерю электролитов (главным образом, натрия). Формирующиеся при этом интег-ративные механизмы регуляции водно-солевого гомеостазиса аналогичны описанным выше при дефиците приема жидкости. При рвоте и поносах происходит потеря жидкости изотоничной плазме крови,
525
т.е. теряется не только вода, но и содержащиеся в ней электролиты, преимущественно натрий и хлориды. Такая дегидратация носит название изотонической и для нее характерен истинный дефицит натрия в организме. Изотоническая дегидратация сопровождается обязательным уменьшением объема циркулирующей плазмы и крови, что определяет формирование приспособительных реакций: повышается нейросекреция вазопрессина за счет рефлекса с волюморецеп-торов предсердий и полых вен (что активирует реабсорбцию воды в почках), усиливается секреция ренина и повышается в крови содержание ангиотензина и альдостерона (что поддерживает сосудистый тонус и вызывает активацию реабсорбции натрия в дистальных канальцах почек). Поскольку при изотонической дегидратации осмотическое давление внеклеточной и внутриклеточной жидкости не меняется, жажда может отсутствовать.
При умеренных физиологических объемах выпиваемой воды (рис. 12.5), еще до поступления ее из желудочно-кишечного тракта в кровь системной циркуляции, снижение осмотического давления
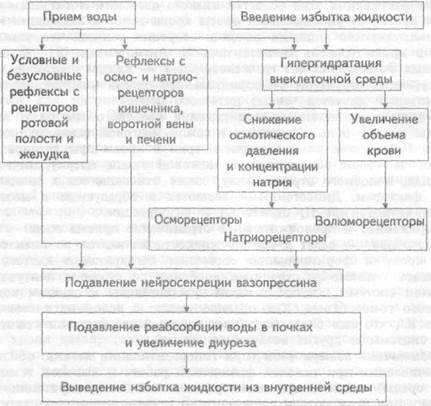
Рис. 12.5. Гомеостатическая регуляция водно-солевого обмена при избытке воды и гипергидратации.
526
 слюны и химуса воспринимаются рецепторами ротовой полости и пищеварительного тракта, а всасывающаяся в кровь воротной вены вода уменьшает концентрацию натрия и осмотическое давление крови поступающей в печень, где этот сдвиг воспринимается соответствующими осмо- и натриорецепторами. Возникающее при их раздражении рефлекторное (в том числе и условнорефлекторное) подавление нейросекреции вазопрессина и нарастание мочеобразо-вания происходят вскоре после приема жидкости. Таким образом, вовлечение систем интегративного контроля водно-солевого баланса начинается еще до сдвигов уровня натрия и осмотического давления в крови системной циркуляции, поэтому в физиологических условиях рефлекторная стимуляция почечной экскреции воды и натрия ограничивает увеличение объема крови из- за всасывания выпитой воды.
слюны и химуса воспринимаются рецепторами ротовой полости и пищеварительного тракта, а всасывающаяся в кровь воротной вены вода уменьшает концентрацию натрия и осмотическое давление крови поступающей в печень, где этот сдвиг воспринимается соответствующими осмо- и натриорецепторами. Возникающее при их раздражении рефлекторное (в том числе и условнорефлекторное) подавление нейросекреции вазопрессина и нарастание мочеобразо-вания происходят вскоре после приема жидкости. Таким образом, вовлечение систем интегративного контроля водно-солевого баланса начинается еще до сдвигов уровня натрия и осмотического давления в крови системной циркуляции, поэтому в физиологических условиях рефлекторная стимуляция почечной экскреции воды и натрия ограничивает увеличение объема крови из- за всасывания выпитой воды.
Избыточный прием воды или введение гипотонических растворов в кровь (рис. 12.5) ведут к гипергидратации, уменьшению осмотического давления и концентрации натрия в плазме. Следствием этого является подавление нейросекреции вазопрессина, в почках стенки дисталъных канальцев и собирательных трубочек остаются непроницаемыми для воды. В то же время, поскольку реабсорбция натрия сохраняется, почки выделяют большие количества гипотоничной мочи и избыток воды удаляется из крови. В отсутствии вазопрессина мочевина не реабсорбируется в собирательных трубочках, что снижает осмолярность интерстиция мозгового вещества почек и тем самым также ослабляет реабсорбцию воды. Если описанные механизмы регуляции не обеспечивают выведение объемов воды, адекватных ее поступившему количеству, устанавливается положительный водный баланс, вода задерживается в организме. Задержка воды в крови (гидремия) снижает ее осмотическое давление и вода выходит в ткани — формируются отеки.
Дата добавления: 2016-03-27; просмотров: 1435;