Водно-солевой обмен.
Водно-солевым обменом называют совокупность процессов поступления воды и электролитов в организм, распределения их во внутренней среде и выделения из организма. У здорового человека поддерживается равенство объемов выделяющейся из организма и поступившей в него за сутки воды, что называют водным балансом организма. Можно рассматривать также и баланс электролитов — натрия, калия, кальция и т.п. Средние показатели водного баланса здорового человека в состоянии покоя показаны в табл. 12.1, а баланса электролитов в табл. 12.2.
При различных возмущающих воздействиях (сдвиги температуры среды, разный уровень физической активности, изменение характера питания) отдельные показатели баланса могут меняться, но сам баланс
495
 Таблица 12.1.
Таблица 12.1.
Средние величины параметров водного баланса организма человека (мл/сут)
| Потребление и образование воды | Выделение воды | ||||||
| Питье и жидкая пища 1200 С мочой 1500 С твердой пищей 1000 С потом 500 Образующаяся эндогенная С выдыхаемым вода "окисления" 300 воздухом 400 | |||||||
| Внутренний цикл жидкостей желудочно-кишечного тракта | |||||||
| Секреция | Реабсорбция | ||||||
| Слюна 1500 Желудочный сок 2500 Желчь 500 Сок pancreas 700 Кишечный сок 3000 | |||||||
| Итого 8200 — 8100 = вода в кале 1 i , | 00 j | ||||||





 Поступление: 2500
Поступление: 2500
Выделение 2500

 Таблица 12.2
Таблица 12.2
Среднесуточный баланс обмена некоторых веществ у человека
| поступление | выделение | ||||
| ЙРШРГТЙЯ | |||||
| пища | метаболизм | моча | фекалии | пот и воздух | |
| Натрий (ммоль) | 2,5 | 2,5 | |||
| Калий (ммоль) | 5,0 | — | |||
| Хлорид (ммоль) | 2,5 | 2,5 | |||
| Азот (г) | — | ||||
| Кислоты (мэкв) | |||||
| нелетучие | - 50 | — | — | ||
| летучие | — | — | |||
 при этом сохраняется. В условиях патологии происходят нарушения баланса с преобладанием либо задержки, либо потерь воды.
при этом сохраняется. В условиях патологии происходят нарушения баланса с преобладанием либо задержки, либо потерь воды.
Вода является важнейшим неорганическим компонентом организма, обеспечивающим связь внешней и внутренней среды, транспорт веществ между клетками и органами. Являясь растворителем органических и неорганических веществ, вода представляет собой основную среду развертывания метаболических процессов. Она входит в состав различных систем органических веществ. Каждый грамм
406
гликогена, например, содержит 1,5 мл воды, каждый грамм белка —
3 мл воды. При ее участии формируются такие структуры как кле
точные мембраны, транспортные частицы крови, макромолекулярные
и надмолекулярные образования. В процессе обмена веществ и
окислении водорода, отделенного от субстрата, образуется эндоген
ная "вода окисления", причем ее количество зависит от вида рас
падающихся субстратов и уровня обмена веществ. Так, в покое при
окислении 100 г жира образуется более 100 мл воды, 100 г белка
— около 40 мл воды, 100 г углеводов — 55 мл воды. Повышение
катаболизма и энергетического обмена ведет к резкому увеличению
образуемой эндогенной воды.
Однако, эндогенной воды у человека недостаточно для обеспечения водной среды метаболических процессов, особенно выведения в растворенном виде продуктов метаболизма. В частности, повышение потребления белков и, соответственно, конечное превращение их в мочевину, удаляемую из организма с мочой, ведет к абсолютной необходимости возрастания потерь воды в почках, что требует повышенного ее поступления в организм. При питании преимущественно углеводной, жировой пищей и небольшом поступлении в организм NaCl потребность организма в поступлении воды меньше. У здорового взрослого человека суточная потребность в воде колеблется от 1 до 3 л.
Общее количество воды в организме составляет у человека от 44 до 70% массы тела или примерно 38-42 л. Содержание ее в разных тканях варьирует от 10% в жировой ткани до 83-90% в почках и крови, с возрастом количество воды в организме уменьшается, также как и при ожирении. У женщин содержание воды ниже, чем у мужчин.
Вода организма образует два водных пространства: внутриклеточное (2/3 обшей воды) и внеклеточное (1/3 общей воды). В условиях патологии появляется третье водное пространство — вода полостей тела: брюшной, плевральной и т.д. Внеклеточное водное пространство включает два сектора: 1) внутрисосудистый водный сектор, т.е. плазму крови, объем которой составляет около 4- 5% массы тела, и 2) интерстициальный водный сектор, содержащий 1/
4 всей воды организма (15% массы тела) и являющийся наиболее
подвижным, меняющим объем при избытке или недостатке воды в
теле. Вся вода организма обновляется примерно через месяц, а
внеклеточное водное пространство — за неделю.
Избыточное поступление и образование воды при неадекватно малом ее выделении из организма ведет к накоплению воды и этот сдвиг водного баланса получил название гипергидратация. При гипергидратации вода накапливается, в основном, в интерстициальном водном секторе. Значительная степень гипергидратации проявляется водной интоксикацией. При этом в интерстициальном водном секторе осмотическое давление становится ниже, чем внутри клеток, они поглощают воду, набухают и осмотическое давление в них становится тоже сниженным. В результате повышенной чувствительности нервных клеток к уменьшению осмолярности водная инток-
497
сикация может сопровождаться возбуждением нервных центров и мышечными судорогами.
Недостаточное поступление и образование воды или чрезмерно большое ее выделение приводят к уменьшению водных пространств, главным образом, интерстициального сектора, что носит название дегидратация. Это сопровождается сгущением крови, ухудшением ее реологических свойств и нарушением гемодинамики. Недостаток в организме воды в объеме 20% массы тела ведет к летальному исходу.
Система регуляции водного баланса обеспечивает два основных гомеостатических процесса: во-первых, поддержание постоянства общего объема жидкости в организме и, во-вторых, оптимальное распределение воды между водными пространствами и секторами организма. К числу факторов поддержания водного гомеостазиса относятся осмотическое и онкотическое давление жидкостей водных пространств, гидростатическое и гидродинамическое давление крови, проницаемость гистогематических барьеров и других мембран, активный транспорт электролитов и неэлектролитов, нейро-эндокринные механизмы регуляции деятельности почек и других органов выделения, а также питьевое поведение и жажда.
Водный баланс организма тесно связан с обменом электролитов. Суммарная концентрация минеральных и других ионов создает определенную величину осмотического давления. Концентрация отдельных минеральных ионов определяет функциональное состояние возбудимых и невозбудимых тканей, а также состояние проницаемости биологических мембран, — поэтому принято говорить о водно-электролитном (или солевом) обмене. Поскольку синтез минеральных ионов в организме не осуществляется, они должны поступать в организм с пищей и питьем. Для поддержания электролитного баланса и, соответственно, жизнедеятельности, организм в сутки должен получать примерно 130 ммоль натрия и хлора, 75 ммоль калия, 26 ммоль фосфора, 20 ммоль кальция и других элементов.
Данные о физиологической роли, суточной потребности и пищевых источниках минеральных ионов приведены в таблице 12.3. В этой же таблице представлены сведения о микроэлементах. К ним относят ту часть минеральных ионов, которые выполняют в организме ряд важных функций, но суточная потребность в этих элементах невелика.
Основным катионом внеклеточного водного пространства является натрий, а анионом — хлор. Во внутриклеточном пространстве основной катион — калий, а анионами являются фосфат и белки.
Для обеспечения физиологических процессов важна не столько общая концентрация каждого электролита в водных пространствах, сколько их активность или эффективная концентрация свободных ионов, поскольку часть ионов находится в связанном состоянии (Са и Mg с протеинами, Na в ячейках клеточных органелл и т.п.). Роль электролитов в жизнедеятельности организма многообразна и неоднозначна. Натрий поддерживает осмотическое давление внекле-
498
Таблица 12.3Физиологическая роль, суточная потребность организма и источники поступления основных минеральных ионов и микроэлементов




 Элементы
Элементы
Физиологическая роль и суточная потребность
Источники




 Натрий Содержится преимущественно во внеклеточ-
Натрий Содержится преимущественно во внеклеточ-
ной жидкости и плазме крови. Играет роль в процессах возбуждения, создании величины осмотического давления жидкостей внутренней среды, распределении и выведении воды из организма; участвует в функции бикарбонатной буферной системы. Суточная потребность 130-155 ммоль.
Кальций Выполняет функцию структурного компонента в тканях зубов и костей, где содержится до 99% общего количества кальция в организме. Вторичный посредник регуляции функций и метаболизма клеток. Необходим для осуществлений процессов возбуждения клеток, синаптической передачи, свертывания крови, сокращения мышц. Суточная потребность 20-30 ммоль.
Калий.Содержится преимущественно внутри клеток,
а также в жидкостях внутренней среды. Необходим для обеспечения возбудимости клеток, проводимости в нервных волокнах, сократимости мышц, основных функциональных свойств миокарда. Суточная потребность 55-80 ммоль.
Хлор Содержится во внеклеточной и внутрикле-
точной жидкостях внутренней среды. Играет роль в процессах возбуждения и торможения, в проведении нервных импульсов, синаптической передаче, образовании соляной кислоты желудочного сока. Суточная потребность 130-155 ммоль.
Фосфор В виде фосфатного аниона содержание в клетках в 40 раз выше, чем во внеклеточной среде. До 80% содержится в костях и зубах в виде минеральных веществ. В составе фосфолипидов входит в структуру клеточных мембран, липопротеидов. Необходимый элемент макроергических соединений и их производных, циклических нуклеотидов, коферментов, играющих важнейшую роль в метаболизме и регуляции физиологических функций. Суточная потребность 20-30 ммоль.
Поваренная соль, растительная и животная пища, жидкости, потребляемые при питье
Молоко и молочные продукты, овощи, зеленые пищевые приправы
Овощи (картофель), мясо, сухофрукты (изюм), орехи
Поваренная соль, растительная и животная пища, жидкости, потребляемые при питье
Молоко, рыба, мясо, яйца, орехи, злаки.
 499
499
Таблица 12.3 (продолжение)
 Железо
Железо
Йод
Магний
Содержится в гемоглобине эритроцитов (до 66%), скелетных мышцах, печени, костном мозге, селезенке. Входит в состав многих ферментов и коферментов. Суточная потребность 170-280 мкмоль.
Необходимый компонент гормонов щитовидной железы, их предшественников и метаболитов. Суточная потребность 1-3 нмоль.
Содержится в костной ткани (необходим для ее образования), скелетных мышцах и нервной системе. Входит в состав многих ферментов и коферментов. Необходим для функции клеточных мембран, сократимости миокарда и гладких мышц. Суточная потребность 10-15 ммоль.
Печень, мясо, рыба, яйца, сухофрукты, орехи.
Морепродукты, рыбий жир, йодированная поваренная
Мясо, молоко, злаки
Медь
Содержится в печени, селезенке, играет Яйца, печень, поч-роль в процессах всасывания железа, син- ки. рыба, шпинат, теза гемоглобина, входит в состав ряда виноград ферментов и пигментов. Суточная потребность 30-80 мкмоль.
Фтор
Сера
Цинк
Содержится в зубных тканях и необходим для их целостности. Входит в состав некоторых ферментов. Суточная потребность около 50 мкмоль. При пятикратной передозировке токсичен.
Входит в состав аминокислот, белков (инсулин) и витаминов, биологически активных веществ, участвует в обезвреживании эндогенных токсинов в печени. Суточная потребность 30-40 ммоль.
Важный компонент ряда ферментов, гормонов. Необходим для процессов роста. Суточная потребность 150-300 мкмоль.
Пищевые продукты, вода, фторированные зубные пасты и NaCI
Мясо, печень, рыба, яйца
Мясо, бобы, крабы, яичный желток
Кобальт
Входит в состав витамина В-12, необходим Печень для нормального эритропоэза. Содержится в печени, костной ткани. Суточная потребность точно не известна, предположительно 2-4 мкмоль.
 точной жидкости, причем его дефицит не может быть восполнен другими катионами. Изменение уровня натрия в жидкостях организма неизбежно влечет за собой сдвиг осмотического давления и в результате — объема жидкостей. Уменьшение концентрации на-
точной жидкости, причем его дефицит не может быть восполнен другими катионами. Изменение уровня натрия в жидкостях организма неизбежно влечет за собой сдвиг осмотического давления и в результате — объема жидкостей. Уменьшение концентрации на-
500
трия во внеклеточной жидкости способствует перемещению воды в клетки, а увеличение содержания натрия вызывает выход воды из клеток. Содержание натрия в клеточной микросреде определяет величину мембранного потенциала и, соответственно, — возбудимость клеток.
Основное количество калия (98%) находится внутри клеток в виде непрочных соединений с белками, углеводами и фосфором. Часть калия содержится в клетках в ионизированном виде и обеспечивает мембранный потенциал. Во внеклеточной среде небольшой количество калия находится преимущественно в ионизированном виде. Обычно выход калия из клеток зависит от увеличения их биологической активности, распада белка и гликогена, недостатка кислорода. Концентрация калия увеличивается при ацидозе и снижается при алкалозе.
Уровень калия в клетках и внеклеточной среде играет важнейшую роль в деятельности сердечно-сосудистой, мышечной и нервной систем, в секреторной и моторной функциях пищеварительного тракта, экскреторной функции почек.
Кальций участвует в физиологических процессах только в ионизированном виде. Этот катион необходим для обеспечения возбудимости нервно- мышечной системы, проницаемости мембран, свертывания крови. Ионизация кальция в крови зависит от рН. При ацидозе содержание ионизированного кальция повышается, а при алкалозе — падает. Алкалоз и снижение уровня кальция ведут к резкому повышению нейромышечной возбудимости и тетании. Влияет на уровень кальция и концентрация белков в плазме крови. Содержание кальция в крови поддерживается в норме в диапазоне 2-4 ммоль/л. Внутриклеточный ионизированный кальций является важнейшим вторичным посредником нервно-гуморальных регуляторных влияний, обеспечивает процессы освобождения медиаторов и секрецию гормонов, энергетику клетки. Основное депо кальция — костная ткань, в которой содержится 90% катиона в связанном виде.
Магний, как и калий, является основным внутриклеточным катионом, т.к. его концентрация в клетках значительно выше, чем во внеклеточной среде. Половина всего магния находится в костях, 49% в клетках мягких тканей и лишь 1% во внеклеточном водном пространстве. Уровень магния в крови составляет 0,7-1 ммоль/л, при этом более 60% катиона находится в ионизированном виде. Магний входит в состав более 300 разных ферментных комплексов, обеспечивая их активность. Он способствует синтезу белков, необходим для поддержания состояния клеточных мембран. Катион уменьшает возбудимость нервно-мышечной системы, сократительную способность миокарда и гладких мышц сосудов, оказывает депрессивное действие на психические функции.
Главным анионом внеклеточной жидкости является хлор, 65% которого находится в ее мобильной части. Концентрация хлора в плазме крови в норме колеблется от 90 до 105 ммоль/л. Специальной физиологической роли этот анион не выполняет, хотя участвует в формировании потенциала покоя возбудимых клеток. Избыток
501
хлора ведет к ацидозу. Анион необходим для образования соляной кислоты в желудке.
Фосфаты являются основными внутриклеточными анионами, где концентрация фосфата выше, чем во внеклеточной среде в 40 раз. Содержание неорганического фосфата в крови составляет 0,94-1,44 ммолль/л, но 50% неорганического фосфата находится в костях, где он вместе с кальцием образуют основное минеральное вещество костной ткани. Фосфаты — необходимый компонент клеточных мембран, играют ключевую роль в метаболических процессах, входя в состав многих коферментов, нуклеиновых кислот и фосфопротеинов, вторичных посредников и макроэргических соединений.
Сульфаты в большем количестве содержатся во внутриклеточном пространстве, входят в состав многих биологически активных веществ. В плазме крови неорганических сульфатов содержится 0,3-1,5 ммоль/л. Они необходимы для обезвреживания токсических соединений в печени.
Для гомеостаза электролитов необходимо взаимодействие нескольких процессов: поступление в организм, перераспределение и депонирование в клетках и их микроокружении, выделение из организма. Поступление в организм зависит от состава и свойств пищевых продуктов и воды, особенностей их всасывания в желудочно-кишечном тракте и состояния энтерального барьера. Однако, несмотря на широкие колебания количества и состава пищевых веществ и воды, водно-солевой баланс в здоровом организме неуклонно поддерживается за счет изменений экскреции с помощью органов выделения. Основную роль в этом гомеостатическом регулировании выполняют почки.
Регуляция водно-солевого обмена, как и большинство физиологических регуляций, включает афферентное, центральное и эфферентное звенья. Афферентное звено представлено массой рецепторных аппаратов сосудистого русла, тканей и органов, воспринимающих сдвиги осмотического давления, объема жидкостей и их ионного состава. В результате, в центральной нервной системе создается интегрированная картина состояния водно-солевого баланса в организме. Следствием центрального анализа является изменение питьевого и пищевого поведения, перестройка работы желудочно-кишечного тракта и системы выделения (прежде всего функции почек), реализуемая через эфферентные звенья регуляции. Последние представлены нервными и, в большей мере, гормональными влияниями.
Функции почек
Функции почек многообразны, при этом часть из них связана с процессами выделения, в которых почки играют ведущую роль, другая же часть может быть названа невыделительными функциями почек. Почки участвуют в регуляции:
1) водного баланса организма (табл. 12.1) и, соответственно, объемов вне- и внутриклеточных водных пространств, поскольку меняют количество выводимой с мочой воды;
502
2) ионного баланса и состава жидкостей внутренней среды путем
избирательного изменения экскреции ионов с мочой;
3) постоянства осмотического давления жидкостей внутренней
среды, за счет изменения количества выводимых осмотически ак
тивных веществ (солей, мочевины, глюкозы и др.);
4) кислотно-основного баланса, путем изменения экскреции во
дородных ионов, нелетучих кислот и оснований (глава 13).
5) метаболизма белков, липидов, углеводов, нуклеиновых кислот и
других органических соединений, во-первых, за счет изменений
экскреции продуктов метаболизма и избытка соединений, поступив
ших с пищей или образовавшихся в организме, во-вторых, благо
даря собственной метаболической функции (синтез аммиака и мо
чевины, новообразование глюкозы, гидролиз белков и липидов, син
тез ферментов, простаноидов и т.п.);
6) циркуляторного гомеостазиса, путем регуляции обмена электро
литов, объема циркулирующей крови, внутренней секреции гормо
нов, регулирующих функции сердечно-сосудистой системы —ренина,
кальцитриола и др. (глава 5), а также экскретируя другие гумораль
ные регуляторы системы кровообращения;
7) эритропоэза, за счет внутренней секреции эритропоэтина —
гуморального регулятора эритрона (глава 6);
8) гемостаза, путем образования гуморальных регуляторов сверты
вания крови и фибринолиза (урокиназы, тромбопластина, тромбок-
сана и простациклина) и участвуя в обмене физиологических анти
коагулянтов (гепарина).
Экскретируя из внутренней среды чужеродные и вредные вещества, почки выполняют защитную функцию. Таким образом, выделяют следующие функции почек: экскреторную, гомеостатическую, метаболическую, инкреторную и защитную. Основной функцией почек, обеспечивающей ведущую роль в выделительной системе организма, является образование и выделение мочи.
Механизмы мочеообразования.Моча образуется в почках из крови, причем почка относится к наиболее интенсивно кровоснабжаемым органам — ежеминутно через почку проходит 1/4 всего объема крови, выбрасываемой сердцем. Основной структурно-функциональной единицей почки, обеспечивающей образование мочи, является нефрон. В почке человека и многих млекопитающих содержится около 1,2 миллионов нефронов. Однако, не все нефроны работают в почке одновременно, существует определенная периодичность функционирования отдельных нефронов, когда часть из них функционирует, а другие нет. Эта периодичность обеспечивает надежность деятельности почки за счет функционального дублирования. В связи с этим важным показателем функциональной активности почки является масса действующих нефронов в конкретный момент времени.
Нефрон состоит из нескольких последовательно соединенных отделов (рис. 12.1), располагающихся в корковом и мозговом веществе почки.
503
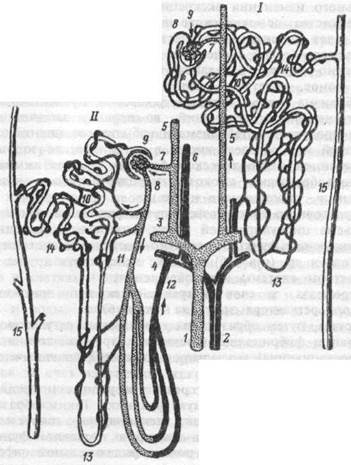
Рис.12.1. Морфологические особенности интракортикальных (I) и юкстамедуллярных (II) нефронов.
I — междолевая артерия, 2 — междолевая вена,
3 — дугообразная венула, 5 — междольковая артериола, 6 — междольковая венула, 7 — приносящая артериаола, 8 — выносящая артериола, 9 — сосудистый клубочек, 10 — проксимальный извитой каналец,
11 — прямой нисходящий сосуд, 12 — прямой восходящий сосуд,
13 — петля Генле, 14 — дистальный извитой каналец,
15 — собирательная трубочка.
1) Сосудистый клубочек или мальпигиевое тельце, находится в корковом веществе, имеет около 50 капиллярных петель, связанных друг с другом и подвешенных как на брыжейке с помощью мезангия, состоящего из волокнистых структур и мезангиальных клеток. Снаружи клубочки покрыты двухслойной капсулой Боумена- Шумлянского. Висцеральный листок капсулы покрывает капилляры клу-
504
бочка и состоит из эпителиальных отростчатых клеток — подоцитов. Отростки подоцитов (большие и малые), называемые педикулами, покрывают всю поверхность капилляров, тесно переплетаясь друг с другом и оставляя межпедикулярные пространства не более 30 нм. Пространства заполнены фибриллярными структурами, образующими щелевую диафрагму, формирующую решетку или сито с диаметром пор около 10 нм. Наружный или париетальный листок капсулы состоит из базальной мембраны, покрытой кубическими эпителиальными клетками, переходящими в эпителий канальцев. Между двумя листками капсулы, расположенными наподобие чаши, имеется щель или полость капсулы, переходящая в просвет главного или проксимального отдела канальцев.
2) Главный или проксимальный отдел канальцев, начинающийся
от полости капсулы извитой частью, которая затем переходит в
прямую часть канальца. Клетки проксимального отдела на апикаль
ной мембране имеют щеточную каемку из микроворсин, покрытых гликокаликсом. Проксимальный отдел расположен в корковом веществе, где переходит в петлю Генле.
3) Тонкий нисходящий отдел петли Генле, который покрыт плос
кими клетками со щелевидными пространствами в цитоплазме ши
риной до 7 нм, спускающийся в мозговое вещество почки, где
поворачивает на 180° и переходит в восходящую часть, являющуюся началом дистального отдела канальцев.
4) Дистальный отдел канальцев, состоящий из восходящей части
петли Генле или прямого отдела и извитой части. Восходящая часть покрыта клетками, напоминающими клетки проксимального отдела, но лишенными щеточной каемки. Извитая часть дистального канальца вновь располагается в коре почки, подходит к клубочку и
обязательно соприкасается с его полюсом между приносящей и
выносящей артериолами. Здесь эпителий канальца становится ци
линдрическим, ядра клеток гиперхромными, этот участок выглядит
темным, плотным, что и дало ему название macula densa — плотное
пятно. Поскольку непрерывная базальная мембрана здесь отсутствует
и клетки эпителия канальца имеют тесный контакт с гранулирован
ными миоэпителиоидными клетками артериолы клубочка, плотное
пятно относят к юкстагломеруллярному аппарату почки. Дистальные
извитые канальцы через короткий связующий отдел впадают в коре
почек в следующий отдел нефрона — собирательные трубки.
5) Собирательные трубки спускаются из коры почек вглубь моз
гового вещества, где их эпителий из кубического становится ци
линдрическим. Темные цилиндрические эпителиальные клетки дис-
тальных отделов собирательных трубок богаты карбангидразой и
обеспечивают секрецию ионов водорода. В глубине мозгового веще
ства в области вершин пирамид собирательные трубки сливаются в
выводные протоки, открывающиеся в полость лоханки.
По особенностям локализации клубочков в коре почек, строения канальцев и особенностям кровоснабжения различают три типа неф-ронов: суперфициальные, интракортикальные и юкстамедуллярные (рис. 12.1).
505
Суперфициальные нефроны имеют поверхностно расположенные в коре клубочки, наиболее короткую петлю Генле, их 20-30%. Интра-кортикальные нефроны, клубочки которых расположены в средней части коры почки, наиболее многочисленны (60-70%) и выполняют основную роль в процессах ультрафильтрации мочи. Диаметр их приносящей артериолы больше, чем у выносящей, ветви последней дают густую сеть капилляров в корковом и мозговом веществе. Юкстамедуллярных нефронов значительно меньше (10-15%), клубочки их расположены у границы коркового и мозгового вещества почки, выносящие артериолы шире приносящих, петли Генле самые длинные и спускаются почти до вершины сосочка пирамид. Выносящие артериолы образуют прямые капиллярные нисходящие и восходящие сосуды, идущие в глубину мозгового вещества параллельно петлям Генле. Юкстамедуллярные нефроны играют ведущую роль в процессах концентрирования и разведения мочи.
Механизм мочеобразования складывается из трех основных процессов: 1) клубочковой ультрафильтрации из плазмы крови воды и низкомолекулярных компонентов с образованием первичной мочи; 2) канальцевой реабсорбции (обратного всасывания в кровь) воды и необходимых для организма веществ из первичной мочи; 3) канальцевой секреции ионов, органических веществ эндогенной и экзогенной природы.
Клубочковая ультрафильтрация и ее регуляция. Процесс клубочковой ультрафильтрации (далее просто фильтрация) осуществляется под влиянием физико-химических и биологических факторов через структуры гломерулярного фильтра, находящегося на пути выхода жидкости из просвета капилляров клубочка в полость капсулы. Гломерулярный фильтр состоит из 3-х слоев: эндотелия капилляров, базальной мембраны и эпителия висцерального листка капсулы или подоцитов. Эндотелий капилляров пронизан отверстиями диаметром до 100 нм, что позволяет свободно проходить через них воде с растворенными в ней веществами, но не форменным элементам крови. На поверхности эндотелия находится особая выстилка — гликокаликс, мешающая доступу форменных элементов и крупных молекул к лежащей под эндотелием базальной мембране. Базалъная мембрана является основной частью фильтра, препятствующей проникновению из плазмы крови крупномолекулярных соединений (белков). При этом не только размер пор мембраны (около 2,9 нм), но и отрицательный заряд препятствуют прохождению молекул с отрицательным зарядом, например альбуминов. Базальная мембрана довольно быстро "изнашивается" и ее элементы непрерывно восстанавливаются с помощью мезангиальных клеток, при этом в течение года происходит полная замена ее основного вещества. Третий слой фильтра образован отростками подоцитов, между которыми остаются щелевые диафрагмы с диаметром пор около 10 нм, поры покрыты гликокаликсом, оставляющим отверстия радиусом около 3 нм. Эта часть фильтра также несет отрицательный заряд.
Поскольку подоциты содержат внутри отростков — педикул акто-миозиновые миофибриллы, они могут сокращаться и расслабляться,
506
действуя как микронасосы, откачивающие фильтрат в полость капсулы. Эта активность подоцитов составляет один из биологических факторов обеспечения процесса фильтрации, к числу которых относится также сокращение и расслабление мезангиалъных клеток, изменяющих тем самым площадь поверхности клубочкового фильтра.
Физико-химические факторы обеспечения фильтрации представлены отрицательным зарядом структур фильтра и фильтрационным давлением, являющимся основной причиной фильтрационного процесса.
Фильтрационное давление — это сила, обеспечивающая движение жидкости с растворенными в ней веществами из плазмы крови капилляров клубочка в просвет капсулы. Эта сила создается гидростатическим давлением крови в капилляре клубочка. Препятствующими фильтрации силами являются онкотическое давление белков плазмы крови (т.к. белки почти не проходят через фильтр) и давление жидкости (первичной мочи) в полости капсулы клубочка. Таким образом, фильтрационное давление (ФД) представляет собой разность между гидростатическим давлением крови в капиллярах (Рг) и суммой онкотического давления плазмы крови (Ро) и давления первичной мочи (Рм) в капсуле: ФД = Рг -(Ро + Рм). Гидростатическое давление крови в капиллярах клубочка высокое, примерно 65-70 мм рт.ст., т.е. почти в 2 раза выше, чем в капиллярах других тканей. Это связано, во-первых, с тем, что капилляры клубочка находятся близко к аорте (короткие почечные и внутрипочечные артерии), и, во-вторых, — диаметр приносящих артериол клубочка больше, чем у выносящих. Гидростатическое давление изменяется при сдвигах соотношения диаметров приносящей и выносящей артериол, что является ведущим механизмом регуляции процесса фильтрации. Онкотическое давление белков плазмы крови составляет около 25-30 мм рт.ст., а давление первичной мочи в капсуле — примерно 15-20 мм рт.ст. Таким образом, ФД составляет в среднем: 70 - (30+20) = 20 мм рт.ст.
Основной количественной характеристикой процесса фильтрации является скорость клубочковой фильтрации (СКФ). СКФ — это объем ультрафильтрата или первичной мочи, образующийся в почках за единицу времени. Эта величина зависит от нескольких факторов: 1) от объема крови, точнее плазмы, проходящей через кору почек в единицу времени, т.е. почечного плазмотока, составляющего в среднем у здорового человека массой 70 кг около 600 мл в мин; 2) фильтрационного давления, обеспечивающего сам процесс фильтрации; 3) фильтрационной поверхности, которая равна примерно 2-3% от общей поверхности капилляров клубочка (1,6 м) и может меняться при сокращении подоцитов и мезангиальных клеток; 4) массы действующих нефронов, т.е. числа клубочков, осуществляющих процесс фильтрации в определенное время.
СКФ поддерживается в физиологических условиях на довольно постоянном уровне (несмотря на изменения системного артериального давления) за счет механизмов ауторегуляции. К их числу относятся: 1) миогенная ауторегуляция тонуса приносящих артериол
507
по принципу феномена Бейлиса-Остроумова (см.главу 7); 2) изменение соотношения тонуса приносящих и выносящих артериол клубочка; 3) активация внутрипочечных гуморальных факторов регуляции почечного кровообращения (ренин-ангиотензинной системы, кининов, простагландинов); 4) изменения числа функционирующих нефронов. Первые два механизма поддерживают постоянство кровотока в клубочках и фильтрационное давление; третий — кроме этого, меняет площадь фильтрационной поверхности и функции подоцитов, четвертый — определяет конечный суммарный эффект ауторегуляции СКФ в органе.
СКФ определяют в результате сопоставления концентрации определенного вещества в плазме крови и моче. При этом, используемое вещество должно выделяться вместе с водой только путем фильтрации и не всасываться в нефроне обратно в кровь. Таким условиям больше всего соответствует полисахарид фруктозы инулин. Исходя из концентрации инулина в плазме [Пин], и, определив его концентрацию в определенном объеме (V) конечной мочи [Мин], рассчитывают какой объем первичной мочи соответствует найденной концентрации инулина. Насколько выросла концентрация инулина в конечной моче по сравнению с его концентрацией в плазме, во столько раз больше объем профильтровавшейся плазмы (т.е. первичной мочи) объема конечной мочи. Этот показатель получил название "клиренса" инулина или коэффициента очищения и рассчитывается по формуле:
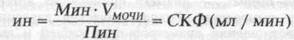
показывающей какой объем плазмы крови в единицу времени вывел найденное количество инулина в мочу или "очистился" от инулина. По мере прохождения мочи по канальцам вода всасывается обратно в кровь и концентрация инулина растет, что и находят в конечной моче.
Поскольку инулин в организме отсутствует, для определения СКФ его необходимо капельно вводить в кровоток, создавая постоянную концентрацию. Это затрудняет исследование, поэтому в клинике обычно используют эндогенное вещество креатинин, концентрация которого в крови довольно стабильна. Клиренс эндогенного креатинина получил название пробы Реберга. Сравнивая клиренс инулина с клиренсом других веществ, определяют процессы, участвующие в выделении этих веществ с мочой. Если клиренс определенного вещества равен клиренсу инулина, значит вещество выделяется почками только путем фильтрации в клубочках. Если клиренс вещества больше клиренса инулина, следовательно, вещество выделяется не только за счет фильтрации, но и секрецией эпителием канальцев. Если клиренс вещества меньше, чем у инулина, — вещество после фильтрации реабсорбируется в канальцах.
В норме СКФ составляет у мужчин около 125 мл/мин, а у женщин — 11О мл/мин. В сутки образуется около 180 л первичной мочи, а за 25 мин фильтруется примерно 3 л плазмы крови, т.е.
508
весь циркулирующий ее объем. За сутки этот объем плазмы крови фильтруется, т.е. очищается, примерно 60 раз. Так как объем конечной мочи около 1,5 л в сутки, очевидно, что из объема первичной мочи за это время всасывается в канальцах обратно в кровь примерно 178,5 л жидкости.
Поскольку первичная моча (клубочковый ультрафильтрат) образуется из плазмы крови, по своему составу она близка плазме, почти полностью лишенной белков. Так, в ультрафильтрате такое же как в плазме крови количество аминокислот, глюкозы, мочевины, креатинина, свободных ионов и низкомолекулярных комплексов. В связи с тем, что белки-анионы не проникают через клубочковый фильтр, для сохранения мембранного равновесия Доннана (равенства произведений концентрации противоположно заряженных ионов электролитов, находящихся по обе стороны мембраны) в первичной моче оказывается на 5% больше концентрация анионов хлора и бикарбоната и, пропорционально меньше концентрация катионов натрия и калия. В первичную мочу проходит небольшое количество наиболее мелких молекул белка —менее 3% гемоглобина и 0,01% альбуминов.
Регуляция СКФ осуществляется за счет нервных и гуморальных влияний. Независимо от природы, регулирующие факторы влияют на СКФ за счет изменения: 1) тонуса артериол клубочков и, соответственно, объемного кровотока (плазмотока) через них и величины фильтрационного давления; 2) тонуса мезангиальных клеток и фильтрационной поверхности; 3) активности подоцитов и их "отсасывающей" функции. Нервные влияния реализуются вазомоторными ветвями почечных нервов, преимущественно симпатической природы, обеспечивающими изменение соотношения тонуса приносящих и выносящих артериол клубочков. Кроме того, симпатические влияния на юкстагломерулярные клетки через бета-адренорецепторы стимулируют секрецию ренина и тем самым реализуют ангиотензинный механизм регуляции фильтрации (спазм выносящих и(или) приносящих артериол). Гуморальные факторы (табл. 12.4) могут как увеличивать, так и уменьшать клубочковую фильтрацию через три описанных выше механизма, причем эффекты вазопрессина реализуются через V-1-рецепторы (глава 5). Важнейшую роль играет ауторегуляция коркового кровотока в почке.
Канальцевая реабсорбция и ее регуляция. Сравнение состава и количества первичной и конечной мочи показывает, что в канальцах нефрона происходит процесс обратного всасывания воды и веществ, профильтровавшихся в клубочках. Этот процесс называется каналъцевой реабсорбцией и в зависимости от отдела канальцев, где он происходит, различают реабсорбцию проксимальную и дистальную. Реабсорбция представляет собой транспорт веществ из мочи в лимфу и кровь и в зависимости от механизма транспорта выделяют пассивную, первично и вторично активную реабсорбцию.
Проксимальная реабсорбция обеспечивает полное всасывание ряда веществ первичной мочи — глюкозы, белка, аминокислот и вита-
509
Таблица 12.4
Основные гормональные влияния на процессы мочеобразования
| Процессы | Увеличивают | Уменьшают |
| Клубочковая фильтрация | Простагландины | Вазопрессин |
| Атриопептид | Ангиотензин-Н | |
| Прогестерон | Норадреналин | |
| Глюкокортикоиды | Адреналин | |
| Окситоцин | Лейкотриены | |
| Глюкагон | ||
| Т-3 и Т-4 | ||
| Паратирин | ||
| Хорионический | ||
| гонадотропин | ||
| Канальцевая реабсорбция | Вазопрессин | Простагландины |
| воды | Пролактин | Атриопептид |
| Ангиотензин-II | Кинины | |
| Инсулин | Паратирин | |
| Эстрогены | Кальцитриол | |
| Хорионический | Т-3 и Т-4 | |
| гонадотропин | Эпифизарный | |
| экстракт |
минов. Впроксимальных отделах всасывается 2/3 профильтровавшихся воды и натрия, большие количества калия, двухвалентных катионов, хлора, бикарбоната, фосфата, а также мочевая кислота и мочевина. К концу проксимального отдела в его просвете остается только 1/3 объема ультрафильтрата, и, хотя его состав уже существенно отличается от плазмы крови, осмотическое давление первичной мочи остается таким же, как в плазме.
Всасывание воды происходит пассивно, по градиенту осмотического давления и зависит от реабсорбции натрия и хлорида. Реабсорбция натрия в проксимальном отделе осуществляется как активным, так и пассивным транспортом. В начальном участке канальцев это активный процесс. Хотя натрий входит в клетки эпителия через апикальную мембрану пассивно через натриевые каналы по концентрационному и электрохимическому градиенту, его выведение через базолатеральные мембраны эпителиальных клеток происходит активно с помощью натрий-калиевых насосов, использующих энергию АТФ. Сопровождающим всасывающийся натрий анионом является здесь бикарбонат, а хлориды всасываются плохо. Объем мочи в канальце уменьшается из- за пассивной реабсорбции воды, и концентрация хлоридов в его содержимом растет. В конечных участках проксимальных канальцев межклеточные контакты высоко проницаемы для хлоридов (концентрация которых повысилась) и они пассивно по градиенту всасываются из мочи. Вместе с ними пассивно реабсорбируются натрий и вода. Такой пассивный транспорт одного иона (натрия) вместе с пассивным транспортом другого (хлорида) носит название котранспорта. Таким образом, в проксимальном отделе нефрона существуют два механизма всасывания воды и ионов: 1) активный транспорт натрия с пассивной реабсорбцией
510
бикарбоната и воды, 2) пассивный транспорт хлоридов с пассивной реабсорбцией натрия и воды. Поскольку натрий и другие электролиты всегда всасываются в проксимальных канальцах с осмотически эквивалентным количеством воды, моча в проксимальных отделах нефрона остается изоосмотичной плазме крови.
Проксимальная реабсорбция глюкозы и аминокислот осуществляется с помощью специальных переносчиков щеточной каемки апикальной мембраны эпителиальных клеток. Эти переносчики транспортируют глюкозу или аминокислоту только если одновременно связывают и переносят натрий. Пассивное перемещение натрия по градиенту внутрь клеток ведет к прохождению через мембрану и переносчика с глюкозой или аминокислотой. Для реализации этого процесса необходима низкая концентрация в клетке натрия, создающая градиент концентрации между внешней и внутриклеточной средой, что обеспечивается энергозависимой работой натрий-калиевого насоса базальной мембраны. Поскольку перенос глюкозы или аминокислоты связан с натрием, а его транспорт определяется активным удалением натрия из клетки, такой вид транспорта называют вторично активным или симпортом, т.е. совместным пассивным транспортом одного вещества (глюкоза) из-за активного транспорта другого (натрия) с помощью одного переносчика.
Поскольку для реабсорбции глюкозы необходимо связывание каждой ее молекулы с молекулой переносчика, очевидно, что при избытке глюкозы может произойти полная загрузка всех молекул переносчиков и глюкоза уже не сможет всасываться в кровь. Эта ситуация характеризуется понятием "максимальный канальцевый транспорт вещества", которое отражает максимальную загрузку канальцевых переносчиков при определенной концентрации вещества в первичной моче и, соответственно, в крови. Постепенно повышая содержание глюкозы в крови и тем самым в первичной моче, можно легко обнаружить ту величину ее концентрации, при которой глюкоза появляется в конечной моче и когда ее экскреция начинает линейно зависеть от прироста уровня в крови. Эта концентрация глюкозы в крови и, соответственно, ультрафильтрате свидетельствует о том, что все канальцевые переносчики достигли предела функциональных возможностей и полностью загружены. В это время реабсорбция глюкозы максимальна и составляет от 303 мг/мин у женщин и до 375 мг/мин у мужчин. Величине максимального канальцевого транспорта соответствует более старое понятие "почечный порог выведения".
Почечным порогом выведения называют ту концентрацию вещества в крови и в первичной моче, при которой оно уже не может быть полностью реабсорбировано в канальцах и появляется в конечной моче. Такие вещества, для которых может быть найден порог выведения, т.е. реабсорбирующиеся при низких концентрациях в крови полностью, а при повышенных концентрациях — не полностью, носят название пороговых. Типичным примером является глюкоза, которая полностью всасывается из первичной мочи при концентрациях в плазме крови ниже 10 моль/л, но появляется в конечной
• 511
моче, т.е. полностью не реабсорбируется, при содержании ее в плазме крови выше 10 моль/л. Следовательно, для глюкозы порог выведения составляет 10 моль/л.
Вещества, которые вообще не реабсорбируются в канальцах (инулин, маннитол) или мало реабсорбируются и выделяются пропорционально накоплению в крови (мочевина, сульфаты и др.), называются непороговыми, т.к. для них порога выведения не существует.
Малые количества профильтровавшегося белка практически полностью реабсорбируются в проксимальных канальцах с помощью пиноцитоза. Мелкие белковые молекулы абсорбируются на поверхности апикальной мембраны эпителиальных клеток и поглощаются ими с образованием вакуолей, которые передвигаясь сливаются с лизосомами. Протеолитические ферменты лизосом расщепляют поглощенный белок, после чего низкомолекулярные фрагменты и аминокислоты переносятся в кровь через базолатеральную мембрану клеток.
Дистальная реабсорбция ионов и воды по объему значительно меньше проксимальной. Однако, существенно меняясь под влиянием регулирующих воздействий, она определяет состав конечной мочи и способность почки выделять либо концентрированную, либо разведенную мочу (в зависимости от водного баланса организма). В дистальном отделе нефрона происходит активная реабсорбция натрия. Хотя здесь всасывается всего 10% от профильтровавшегося количества катиона, этот процесс обеспечивает выраженное уменьшение его концентрации в моче и, напротив, повышение концентрации в интерстициальной жидкости, что создает значительный градиент осмотического давления между мочой и интерстицием. Хлор всасывается преимущественно пассивно вслед за натрием. Способность эпителия дистальных канальцев секретировать в мочу Н-ионы связана с реабсорбцией ионов натрия, этот вид транспорта в виде обмена натрия на протон получил название "антипорт". Активно всасывается в дистальном отделе канальцев калий, кальций и фосфаты. В собирательных трубочках, главным образом юкстамедул-лярных нефронов, под влиянием вазопрессина повышается проницаемость стенки для мочевины и она, благодаря высокой концентрации в просвете канальца, пассивно диффундирует в окружающее интерстициальное пространство, увеличивая его осмолярность. Под влиянием вазопрессина стенка дистальных извитых канальцев и собирательных трубочек становится проницаемой и для воды, в результате чего происходит ее реабсорбция по осмотическому градиенту в гиперосмолярный интерстиций мозгового вещества и далее в кровь.
Способность почки образовывать концентрированную или разведенную мочу обеспечивается деятельностью противоточно-множительной канальцевой системы почки, которая представлена параллельно расположенными коленами петли Генле и собирательными трубочками (рис.12.2). Моча двигается в этих канальцах в противоположных направлениях (почему систему и назвали противоточной), а процессы транспорта веществ в одном колене системы усилива-
512
ются ("умножаются") за счет деятельности другого колена. Определяющую роль в работе противоточного механизма играет восходящее колено петли Генле, стенка которого непроницаема для воды, но активно реабсорбирует в окружающее интерстициальное пространство ионы натрия. В результате, интерстициальная жидкость становится гиперосмотичной по отношению к содержимому нисходящего колена петли и по направлению к вершине петли осмотическое давление в окружающей ткани растет. Стенка же нисходящего колена проницаема для воды, которая пассивно уходит из просвета в гиперосмотичный интерстиций. Таким образом, в нисходящем колене моча из-за всасывания воды становится все более и более гиперосмотичной, т.е. устанавливается осмотическое равновесие с интерстициальной жидкостью. В восходящем колене, из-за всасывания натрия, моча становится все менее осмотичной и в корковый отдел дистального канальца восходит уже гипотоничная моча. Однако ее количество из-за всасывания воды и солей в петле Генле существенно уменьшилось.
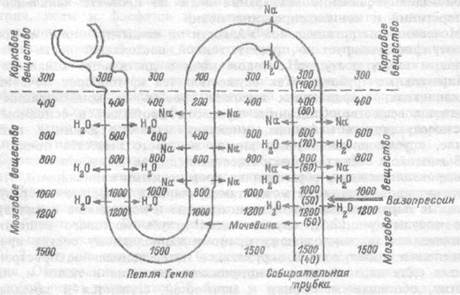
Рис. 12.2. Противоточно- множительная тубулярная система мозгового вещества почки.
Цифрами обозначены величины осмотического давления интерстициальной жидкости и мочи. В собирательной трубочке цифрами в скобках обозначено осмотическое давление мочи в отсутствие вазопрессина (разведение мочи), цифрами без скобок — осмотическое давление мочи в условиях действия вазопрессина (концентрирование мочи).
 513
513
Собирательная трубочка, в которую затем поступает моча, тоже образует с восходящим коленом петли Генле противоточную систему. Стенка собирательной трубочки становится проницаемой для воды только в присутствии вазопрессина. В этом случае, по мере продвижения мочи по собирательным трубочкам вглубь мозгового вещества, в котором нарастает осмотическое давление из-за всасывания натрия в восходящем колене петли Генле, все больше воды пассивно уходит в гиперосмотичный интерстиций и моча становится все более концентрированной.
Под влиянием вазопрессина реализуется еще один важный для концентрирования мочи механизм — пассивный выход мочевины из собирательных трубочек в окружающий интерстиций. Всасывание воды в верхних отделах собирательных трубочек ведет к нарастанию концентрации мочевины в моче, а в самых нижних их отделах, расположенных в глубине мозгового вещества, вазопрессин повышает проницаемость для мочевины и она пассивно диффундирует в интерстиций, резко повышая его осмотическое давление. Таким образом, интерстиций мозгового вещества становится наиболее высоко осмотичным в области вершины почечных пирамид, где и происходит увеличение всасывания воды из просвета канальцев в интерстиций и концентрирование мочи.
Мочевина интерстициальной жидкости по концентрационному градиенту диффундирует в просвет тонкой восходящей части петли Генле и вновь поступает с током мочи в дистальные канальцы и собирательные трубочки. Так осуществляется кругооборот мочевины в канальцах, сохраняющих высокий уровень ее концентрации в мозговом веществе. Описанные процессы протекают в основном в юкстамедуллярных нефронах, имеющих наиболее длинные петли Генле, спускающиеся глубоко внутрь мозгового вещества почки.
В мозговом веществе почки имеется и другая — сосудистая противоточная система, образованная кровеносными капиллярами. Поскольку кровеносная сеть юкстамедуллярных нефронов образует длинные параллельные прямые нисходящие и восходящие капиллярные сосуды (рис. 12.1), спускающиеся вглубь мозгового вещества, двигающаяся по нисходящему прямому капиллярному сосуду кровь постепенно отдает воду в окружающее интерстициальное пространство в силу нарастающего осмотического давления в ткани и, напротив, обогащается натрием и мочевиной, сгущается и замедляет свое движение. В восходящем капиллярном сосуде по мере движения крови в ткани с постепенно снижающимся осмотическим давлением происходят обратные процессы — натрий и мочевина по концентрационному градиенту диффундируют обратно в ткань, а вода всасывается в кровь. Таким образом, и эта противоточная система способствует поддержанию высокого осмотического давления в глубоких слоях ткани мозгового вещества, обеспечивая удаление воды и удержание натрия и мочевины в интерстиций.
Деятельность описанных противоточных систем во многом зависит от скорости движения находящихся в них жидкостей (мочи или крови). Чем скорее будет двигаться моча по трубкам противоточной
514
системы канальцев, тем меньшие количества натрия, мочевины и воды успеют реабсорбироваться в интерстиций и большие количества менее концентрированной мочи будут выделяться почкой. Чем выше будет скорость кровотока по прямым капиллярным сосудам мозгового вещества почки, тем больше натрия и мочевины унесет кровь из почечного интерстиция, т.к. они не успеют диффундировать из крови назад в ткань. Этот эффект называют "вымыванием" осмотически активных веществ из интерстиция, в результате его осмолярность падает, концентрирование мочи уменьшается и почкой выделяется больше мочи низкого удельного веса (разведение мочи). Чем медленнее происходит движение мочи или крови в мозговом веществе почек, тем больше осмотически активных веществ накапливается в интерстиции и выше способность почки концентрировать мочу.
Регуляция каналъцевой реабсорбции осуществляется как нервным, так и, в большей мере, гуморальным путем.
Нервные влияния преимущественно реализуются симпатическими проводниками и медиаторами через бета- адренорецепторы мембран клеток проксимальных и дистальных канальцев. Симпатические эффекты проявляются в виде активации процессов реабсорбции глюкозы, натрия, воды и фосфатов и реализуются через систему вторичных посредников (аденилатциклаза — цАМФ). В регуляции процессов метаболизма почечной ткани существенную роль играют трофические влияния симпатической нервной системы. Нервная регуляция кровообращения в мозговом веществе почки увеличивает или уменьшает эффективность сосудистой противоточной системы и концентрирование мочи. Сосудистые эффекты нервной регуляции могут опосредоваться через внутрипочечные системы гуморальных регуляторов — ренин- ангиотензинную, кининовую, простагландины и др.
Основным фактором регуляции реабсорбции воды в дистальных отделах нефрона является гормон вазопрессин, называвшийся ранее антидиуретическим гормоном. Этот гормон образуется в супраопти-ческом и паравентрикулярных ядрах гипоталамуса и поступает в кровь из нейрогипофиза. Влияние вазопрессина на проницаемость эпителия канальцев обусловлено наличием рецепторов к гормону, относящихся к V-2 типу, на поверхности базолатеральной мембраны клеток эпителия. Образование гормон-рецепторного комплекса (глава 3), влечет за собой через посредство GS-белка и гуанилового нуклеотида активацию аденилатциклазы и образование цАМФ у базолатеральной мембраны (рис. 12.3). После этого цАМФ пересекает клетку эпителия и, достигнув апикальной мембраны, активирует цАМФ- зависимые проте-инкиназы. Под влиянием этих ферментов происходит фосфорилирова-ние мембранных белков, приводящее к повышению проницаемости для воды и увеличению поверхности мембраны. Перестройка ультраструктур клетки ведет к образованию специализированных вакуолей, переносящих большие потоки воды по осмотическому градиенту от апикальной к базолатеральной мембране, не позволяя самой клетке набухать. Такой транспорт воды через клетки эпителия реализуется вазопрессином в собирательных трубочках. Кроме того, в дистальных
515


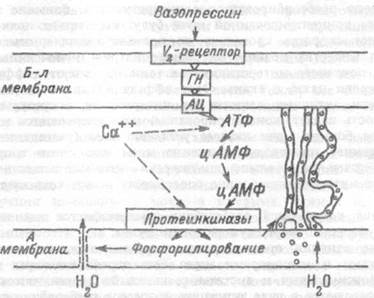
Рис. 12.3. Механизм действия вазопрессина на проницаемость собирательных трубочек для воды.
Б-л мембрана — базолатеральная мембрана клеток,
А мембрана — апикальная мембрана,
ГН — гуанидиновый нуклеотид, АЦ — аденилатциклаза.
 канальцах вазопрессин обусловливает активацию и выход из клеток гиалуронидаз, вызывающих расщепление гликозаминогликанов основного межклеточного вещества и межклеточный пассивный транспорт воды по осмотическому градиенту.
канальцах вазопрессин обусловливает активацию и выход из клеток гиалуронидаз, вызывающих расщепление гликозаминогликанов основного межклеточного вещества и межклеточный пассивный транспорт воды по осмотическому градиенту.
Канальцевая реабсорбция воды регулируется и другими гормонами. С учетом механизмов действия все гормоны, регулирующие реабсорбцию воды, можно представить в виде шести групп:
1) повышающие проницаемость мембран дистальных отделов неф-
рона для воды (вазопрессин, пролактин, хорионический гонадотро-
пин);
2) меняющие чувствительность клеточных рецепторов к вазопрес-
сину (паратирин, кальцитонин, кальцитриол, простагландины, аль-
достерон);
3) меняющие осмотический градиент интерстиция мозгового слоя
почки и, соответственно, пассивный осмотический транспорт воды
(паратирин, кальцитриол, тиреоидные гормоны, инсулин, вазопрессин);
4) меняющие активный транспорт натрия и хлорида, а за счет
этого и пассивный транспорт воды (альдостерон, вазопрессин, ат-
риопептид, прогестерон, глюкагон, кальцитонин, простагландины);
5) повышающие осмотическое давление канальцевой мочи за счет
нереабсорбированных осмотически активных веществ, например глю
козы (контринсулярные гормоны);
516
6) меняющие кровоток по прямым сосудам мозгового .вещества и, тем самым, накопление или "вымывание" осмотически активных веществ из интерстиция (ангиотензин- II, кинины, простагландины, паратирин, вазопрессин, атриопептид).
Канальцевая реабсорбция электролитов, также как и воды, регулируется преимущественно гормональными, а не нервными влияниями.
Реабсорбция натрия в проксимальных канальцах активируется аль-достероном и угнетается паратирином, в толстой части восходящего калена петли Генле реабсорбция натрия активируется вазопресси-ном, глюкагоном, кальцитонином, а угнетается простагландинами Е. В дистальном отделе канальцев основными регуляторами транспорта натрия являются альдостерон (активация), простагландины и атриопептид (угнетение). Регуляция канальцевого транспорта кальция, фосфата и частично магния обеспечивается, в основном, кальций-регулирующими гормонами. Паратирин имеет в канальцевом аппарате почки несколько участков действия. В проксимальных канальцах (прямой отдел) всасывание кальция происходит параллельно с транспортом натрия и воды. Угнетение реабсорбции натрия в этом отделе под влиянием паратирина сопровождается параллельным снижением реабсорбции кальция. За пределами проксимального канальца паратирин избирательно усиливает реабсорбцию кальция, особенно в дистальном извитом канальце и корковой части собирательных трубочек. Реабсорбция кальция активируется также кальцитриолом, а подавляется кальцитонином. Всасывание фосфата в канальцах почки угнетается и паратирином (проксимальная реабсорбция), и кальцитонином (дистальная реабсорбция), а усиливается кальцитриолом и соматотропином. Паратирин активирует реабсорбцию магния в корковой части восходящего колена петли Генле и тормозит проксимальную реабсорбцию бикарбоната.
Канальцевая секреция и ее регуляция. Каналь-цевой секрецией называют активный транспорт в мочу веществ, содержащихся в крови или образуемых в самих клетках канальцевого эпителия, например аммиака. Секреция осуществляется, как правило, против концентрационного или электрохимического градиента с затратами энергии. Путем канальцевой секреции из крови выделяются как ионы (К+, Н+), органические кислоты и основания эндогенного происхождения, так и поступившие в организм чужеродные вещества, в том числе органического происхождения. Для ряда чужеродных организму веществ органической природы (антибиотиков, красителей и рентгеноконтрастных препаратов) скорость и интенсивность выделения из крови путем канальцевой секреции значительно превышает их выведение путем клубочковой фильтрации. Таким образом, канальцевая секреция является одним из механизмов обеспечения гомеостазиса.
Способностью к секреции обладают клетки эпителия и проксимального, и дистальных отделов канальцев. При этом, клетки проксимальных канальцев секретируют органические соединения с помощью специальных переносчиков: один из которых обеспечивает
секрецию органических кислот (парааминогиппуровой кислоты, ди-одраста, фенолрота, пенициллина и др.), а другой — секрецию органических оснований (гуанидина, пиперидина, тиамина, холина, се-ротонина, хинина, морфина и т.п.). Секреция водородных ионов происходит в проксимальных канальцах в большей мере, чем в дистальных. Однако, дистальная секреция водородных ионов играет основную роль в регуляции кислотно-основного состояния внутренней среды (см. главу 13).
Секреция калия происходит в дистальных канальцах и собирательных трубочках, регуляция осуществляется альдостероном, усиливающим секрецию К+ и подавляющим его реабсорбцию. Секреция аммиака, образующегося в самих клетках эпителия, происходит и в проксимальном, и в дистальном отделах.
Регуляция канальцевой секреции осуществляется с помощью гормонов и симпатической нервной системы. Эффекты нервной регуляции осуществляются за счет изменений кровотока в постгломеру-лярных капиллярах почки, т.е. транспорта веществ с кровью секре-тирующим клеткам, и влияния на энергетический обмен в клетках эпителия канальцев. К числу гормонов, усиливающих проксимальную канальцевую секрецию органических веществ за счет метаболических эффектов, относятся соматотропин аденогипофиза, иодсодер-жащие гормоны щитовидной железы и андрогены.
Процесс секреции некоторых веществ в проксимальных канальцах идет настолько интенсивно, что за одно прохождение крови через корковое вещество почек из нее удаляются полн
Дата добавления: 2016-03-27; просмотров: 1954;