НИСХОДЯЩИЙ КОНТРОЛЬ ДЕЯТЕЛЬНОСТИ СПИННОГО МОЗГА
Деятельность спинного мозга не исчерпывается его участием в осуществлении разнообразных рефлекторных реакций. Большое значение имеет также активность спинальных центров, возникающая в результате импульсации из различных вышерасположенных нервных структур, которые играют существенную роль в координации рефлекторной деятельности и в регуляции афферентных систем, доставляющих в мозг информацию от различных рецепторных образований.
Супраспинальные центры посредством нисходящих .путей контролируют активность всех основных нервных элементов спинного мозга: мотонейронов, вставочных нейронов, нейронов восходящих систем, первичных афферентных волокон.
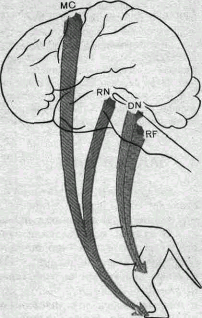 Рис. 84. Моносинаптические проекции различных нисходящих трактов к мононейронам мышц задней конечности обезьяны (схема).
RF — ретикулярная формация; DN — ядро Дейтерса~; RN — красное ядро; МС — моторная зона коры.
Рис. 84. Моносинаптические проекции различных нисходящих трактов к мононейронам мышц задней конечности обезьяны (схема).
RF — ретикулярная формация; DN — ядро Дейтерса~; RN — красное ядро; МС — моторная зона коры.
|
Наиболее толстые, быстропроводящие волокна ретикуло- и вестибулоспинального трактов, а у приматов — рубро- и кортико-спинального трактов способны возбуждать мотонейроны спинного мозга моносинаптически, без переключения на вставочных нейронах. Возбуждающие постсинаптические потенциалы, возникающие в мотонейронах спинного мозга при активации быстропроводящих волокон нисходящих трактов (устанавливающих с ними моносинаптические связи), характеризуются коротким латентным периодом, способностью воспроизводить высокие ритмы раздражения, отсутствием пресинаптического торможения со стороны афферентных систем. Эти свойства обеспечивают высокую эффективность и специфичность нисходящих команд, поступающих по наиболее короткому пути из мозга к мотонейронам.
Волокна каждого тракта активируют преимущественно определенные по функции мотонейроны. Так, моносинаптические кортико-спинальные влияния наиболее активны в мотонейронах, иннервирующих дистальные мышцы конечностей (в первую очередь мышцы пальцев кисти и стопы). Мотонейроны проксимальных мышц конечностей и мышц туловища стимулируются главным образом волокнами ретикуло- и вестибулоспинального трактов. Схема моносинаптических проекций, идущих из разных отделов мозга к мотонейронам задней конечности обезьяны, показана на рис. 84.
Нисходящие тракты оказывают влияние и на различные типы вставочных нейронов спинного мозга и таким образом могут изменять течение его различных рефлекторных реакций.
Способность нисходящих систем контролировать вторичные афферентные нейроны, дающие начало восходящим трактам спинного мозга, отчетливо проявляется в том, что нисходящие импульсы вызывают возбуждение и тормозные синаптические реакции во многих нейронах восходящих трактов. Наконец, импульсация в спинной мозг по волокнам нисходящих путей способна вызывать деполяризацию терминалей первичных афферентных волокон, т. е. пресинаптическое торможение-
Таким образом, импульсы, поступающие в спинной мозг по нисходящим путям, могут воздействовать непосредственно на спинальные двигательные центры, контролировать влияние на мотонейроны через полис инаптичес кие пути различной сложности и даже видоизменять информацию, поступающую в мозг по первичным и вторичным афферентным нейронам.
Такой всеобъемлющий контроль нейронного аппарата спинного мозга супраспинальными структурами обеспечивает значительную зависимость спинальных функций от вышележащих центров. Поэтому частичные и тем более полные перерезки спинного мозга вызывают резкие нарушения деятельности спинальных сегментов каудальнее места перерезки (спинальный шок), в первую очередь из-за выключения связей с головным мозгом.
При односторонних поражениях спинного мозга, возникающих в результате травмы или различного рода заболеваний (опухоль, рассеянный склероз и т. д.), развивается сложный симпто-мокомплекс, известный под названием синдром Броун-Секара. На стороне поражения развиваются паралич, расстройство мышечной и болевой чувствительности, сосудодвигательные нарушения. На противоположной стороне произвольные движения сохраняются, однако болевая и температурная чувствительность исчезает при сохранении мышечной чувствительности. Тактильная чувствительность несколько уменьшается, но не исчезает на обеих сторонах.
Восстановление рефлекторной деятельности развивается постепенно и тем медленнее, чем выше организация нервной системы. Наиболее выражены явления спинального шока у человека. Раньше всего восстанавливается работа двухнейронной рефлекторной дуги — рефлексы растяжения, затем рефлекс сгибания конечности в ответ на болевое раздражение. Рефлекторное опорожнение мочевого пузыря и сосудистые рефлексы восстанавливаются позднее. Наблюдается значительная генерализация спинальных рефлексов вследствие ослабления координации между различными спинальными центрами.
ЗАДНИЙ МОЗГ
Задний мозг, включающий продолговатый мозг и мост мозга (варолиев мост), представляет собой филогенетически древнюю область"ЦНС. Это находит отражение в том, что задний мозг сохраняет определенные черты сегментарного строения.
В заднем мозге локализованы скопления нервных клеток, образующих ядерные структуры, а также проводящие пути, идущие из спинного мозга и из различных образований, расположенных выше заднего мозга. В задний мозг поступают афферентные волокна от вестибулярных и слуховых рецепторов, от кожи и мышц головы, от внутренних органов. Черты сегментарного строения этого отдела мозга проявляются в том, что здесь локализованы ядра V—XII пар черепных нервов, часть из которых подобно мотонейронам спинного мозга, иннервируют лицевую и глазодвигательную мускулатуру.
Нейронная организация заднего мозга по сравнению с таковой спинного мозга отличается большой сложностью. Однако, как и в спинном мозге, здесь можно выделить эфферентные нейроны (включая мотонейроны), вставочные нейроны, нейроны восходящих и нисходящих трактов, первичные афферентные волокна, волокна проводящих путей, идущих через область заднего мозга в восходящем (ростральном) и нисходящем (каудальном) направлении.
Наибольшим сходством со спинальными нейронными центрами обладают ядра черепных нервов, получающие афферентные импульсы с периферии и посылающие эфферентные импульсы к мышцам и другим органам и тканям.
Общая схема строения продолговатого мозга и моста и расположенных в них ядер показана на рис. 85.
Ядра подъязычного (XII пара) и добавочного (XI пара) нервов являются чисто двигательными. Аксоны расположенных в этих ядрах мотонейронов иннервируют соответственно мускулатуру языка и мышцы, осуществляющие движение головы.
Соматомоторные и вегетативные ядра смешанных блуждающего (X пара) и языког лоточного (IX пара) нервов образованы скоплением нервных клеток с менее четким обособлением на отдельные ядерные структуры. Двигательное ядро Х—IX пар черепных нервов расположено в вентролатеральной части продолговатого мозга. Аксоны нейронов двигательных ядер этих нервов идут в составе веточек блуждающего и языкоглоточного нервов и иннервируют мышцы глотки и гортани.
Висцеросенсорное ядро Х—IX пар (называемое ядром солитарного пучка} получает чувствительные волокна от афферентных нейронов, тела которых находятся в яремном, пучковидном и каменистом узлах (которые гомологичны спинномозговым ганглиям). По этим афферентным путям поступают импульсы от рецепторов языка, гортани, трахеи, пищевода, внутренних органов грудной и брюшной полости. Висцеросенсорное ядро связано посредством вставочных нейронов с висцеромоторными ядрами блуждающего и языкоглоточного нервов. Расположенные в этих ядрах эфферентные нейроны иннервируют околоушную железу, железистые и гладкомышечные клетки трахеи, бронхов, желудка и кишечника, а также сердце и сосуды.
Восьмая пара черепных нервов (последняя, располагающаяся в пределах продолговатого мозга) является чувствительной. В составе VIII нерва имеются две ветви — вестибулярная и слуховая.
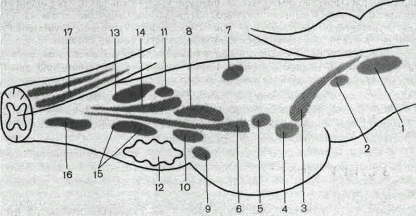
Рис. 85. Продолговатый мозг и мост мозга (варолиев мост) с расположенными в них ядрами черепных нервов (схема).
1 — ядро глазодвигательного нерва; 2 — ядра блокового нерва; 3 — среднемозговое ядро тройничного нерва;
4 — двигательное ядро тройничного нерва; 5 — основное ядро тройничного нерва; 6 — ядро отводящего нерва; 7—8—вестибулярные ядра; 9—кохлеарное ядро; 10—ядро лицевого нерва; 11—парасимпатические ядра языкоглоточного и блуждающего нервов; 12 — олива; 13 — ядро подъязычного нерва; 14 — ядро солитарного тракта; 15—двигательное ядро блуждающего нерва; 16—ядро дополнительного нерва;
17—ядро тонкого пучка (Голля) и клиновидного пучка (ядро Бурдаха).
Слуховая ветвь образована афферентными волокнами, идущими от спирального (кортиева) органа улитки. Слуховые афферентные волокна вступают в продолговатый мозг позади веревчатого тела и достигают вентрального и дорсального слуховых ядер. Аксоны нейронов слуховых ядер поднимаются по боковой петле и достигают соответствующих переключении слуховой системы.
Значительная часть вестибулярных волокон, идущих от рецепторов полукружных каналов, заканчивается на нейронах вестибулярных ядер: медиального (ядро Швальбе), преддверного верхнего (ядро Бехтерева), преддверного латерального (ядро Дейтерса) и нисходящего. Кроме того, часть волокон вестибулярного нерва направляется в мозжечок. Нейроны вестибулярных ядер дают начало вестибуломозжечковому и вестибулоспинальному трактам. Наиболее мощная часть вестибулоспинального пути берет начало от фаланговых наружных клеток (клеток Дейтерса).
В медиальной части продолговатого мозга находится ретикулярная, или сетевидная, формация, представляющая собой скопление нейронов разных размеров и формы, разделенных множеством проходящих в различных направлениях волокон. Это переплетение волокон и клеток, образующее ретикулярную формацию мозгового ствола, продолжается и в мосту мозга (варолиевом мосту) и в среднем мозге.
На уровне продолговатого мозга принято выделять несколько ядер ретикулярной формации:
ретикулярное гигантоклеточное, ретикулярное мелкоклеточное ядро и латеральное ретикулярное.
Клетки ретикулярной формации являются началом как восходящих, так и нисходящих путей, дающих многочисленные коллатерали, окончания которых образуют синаптические контакты на нейронах разных ядер ЦНС.
Волокна ретикулярных клеток, направляющиеся в спинной мозг, образуют ретикулоспинальный тракт.
Нейроны ретикулярной формации продолговатого мозга получают многочисленные коллатерали от волокон восходящих путей, начинающихся в спинном мозге, в том числе от спиноретикулярных волокон, а также от нейронов вышележащих отделов мозга:
из коры больших полушарий, из ядер мозжечка. .
В мосту мозга, являющемся как бы продолжением продолговатого мозга, связывающим последний со средним мозгом, расположены ядра трех следующих пар черепных нервов: лицевого (VII пара), отводящего (VI пара) и тройничного (V пара). Лицевой нерв является смешанным. Идущие в его составе афферентные волокна передают сигналы от вкусовых рецепторов передней части языка. Эфферентные волокна лицевого нерва иннервируют мимическую мускулатуру лица.
Отводящий нерв является двигательным. Его мотонейроны, расположенные на дне IV желудочка в задней половине моста, иннервируют наружную прямую мышцу глаза.
Тройничный нерв, так же как и лицевой, является смешанным. В его двигательном ядре находятся клетки, иннервирующие жевательные мышцы, мышцы небной занавески и мышцу, напрягающую барабанную перепонку. Чувствительное ядро тройничного нерва, начинаясь еще в каудальном конце продолговатого мозга, простирается через весь мост, вплоть до рострального конца среднего мозга.
К клеткам этого ядра подходят аксоны афферентных нейронов полулунного ганглия, доставляющие сигналы от рецепторов кожи лица, теменной и височной области, конъюнктивы, слизистой оболочки носа, надкостницы костей черепа, зубов, твердой мозговой оболочки, языка.
Ядра ретикулярной формации моста по существу являются продолжением ядер ретикулярной формации продолговатого мозга. Наиболее крупное из них — каудальное ретикулярное ядро моста — можно рассматривать как продолжение ретикулярного гигантоклеточного ядра продолговатого мозга. Клетки его (так же, как и нейроны гигантоклеточного ядра продолговатого мозга) дают начало волокнам ретикулоспинального тракта. Кроме того, можно выделить оральное ретикулярное ядро моста, переходящее в клеточную группу, образующую ретикулярную формацию среднего мозга, и ретикулярное ядро покрышки моста. Аксоны нервных клеток покрышки моста проецируются в мозжечок. Из медиальных ядер ретикулярной формации моста мозга начинаются восходящие волокна, направляющиеся в средний и промежуточный мозг.
Дата добавления: 2016-03-27; просмотров: 3476;