МЕХАНИЗМЫ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ
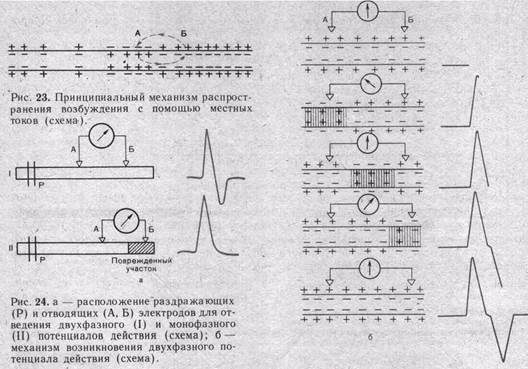
Проведение возбуждения вдоль нервных и мышечных волокон осуществляется при помощи так называемых местных токов, возникающих между возбужденным (деполяризованным) и покоящимися (нормально поляризованными) участками волокна. Распространение местных токов по длине волокна определяется его кабельными свойствами. Направление местного тока, показанного на рис. 23 таково, что он деполяризует соседний с активным (А) покоящийся (В) участок мембраны. Деполяризация эта быстро достигает критического уровня и порождает потенциал действия, который в свою очередь активирует соседний покоящийся участок. Благодаря такому эстафетному механизму возбуждение распространяется вдоль всего волокна. В мышечных и без-
мякотных нервных волокнах возбуждение осуществляется непрерывно «от точки к точке». Особенности проведения возбуждения по миелинизированным волокнам рассмотрены далее.
Распространение возбуждения в нерве или мышце можно зарегистрировать в эксперименте, если к двум точкам —А и Б (рис. 24, а) приложить отводящие электроды, связанные с регистрирующей аппаратурой, а к другой точке (Р) — раздражающие электроды. При нанесении электрического стимула на экране осциллографа регистрируется двухфазное колебание потенциала.
Схема на рис. 24, б разъясняет механизм возникновения этих двух противоположно направленных отклонений потенциала. В состоянии покоя все участки наружной поверхности возбудимой мембраны заряжены электроположительно по отношению к ее внутренней поверхности. Когда волна возбуждения проходит через участок под электродом, ближайшим к месту раздражения, наружная поверхность мембраны в этом участке становится электроотрицательной по отношению к точке Б. Это вызывает отклонение луча осциллографа вверх. Когда волна возбуждения покидает этот участок, луч возвращается в исходное положение. Затем возбуждение достигает участка под вторым электродом Б; этот участок в свою очередь становится электроотрицательным по отношению к точке А, а луч осциллографа отклоняется вниз. Если участок нерва под дальним электродом Б сделать невозбудимым при воздействии какого-либо агента, например новокаина, либо нарушить проведение возбуждения между участками А и Б, вторая фаза колебаний потенциала исчезает и регистрируемый потенциал действия становится однофазным.
Теория проведения возбуждения при помощи местных токов впервые была выдвинута Германом в 1899 г. В настоящее время она получила подтверждение в большом числе экспериментов. Так, показано, что если участок нервного волокна поместить в среду, лишенную ионов и, следовательно, обладающую очень высоким сопротивлением (такой средой может быть, например, раствор сахарозы), то проведение возбуждения через этот участок полностью прекратится. Однако оно тотчас восстанавливается, если два разобщенных неэлектролитом участка волокна соединить металлическим проводником.
Скорость проведения зависит не только от сопротивления окружающей волокно среды, но.и от внутреннего сопротивления волокна (т. е. сопротивления аксоплазмы на единицу длины). С увеличением диаметра волокна это сопротивление падает, поэтому скорость проведения возрастает. При одном и том же диаметре волокна скорость зависит главным образом от величины так называемого фактора надежности, который представляет собой отношение:

Чем фактор надежности больше, тем скорость проведения выше и наоборот. В нервных волокнах фактор надежности обычно 5—6. Это означает, что для покоящихся участков мембраны распространяющийся потенциал действия является сильным раздражителем, обладающим большим избытком мощности. Поэтому, для того чтобы заблокировать проведение нервного импульса, необходимо либо сильно повысить величину порога деполяризации нервного волокна, либо очень значительно снизить амплитуду его потенциала действия. Местные обезболивающие препараты (новокаин, кокаин, дикаин), применяемые в медицинской практике, вызывают оба этих изменения одновременно.
ОБМЕН ВЕЩЕСТВ ПРИ ВОЗБУЖДЕНИИ
При возникновении и проведении возбуждения в нервных клетках и мышечных волокнах происходит усиление обмена веществ. Это проявляется как рядом биохимических изменений, происходящих в мембране и цитоплазме клеток, так и усилением их теплопродукции.
Биохимическими и гистохимическими методами исследования установлено, что при возбуждении наблюдается усиление распада в клетках богатых энергией фосфорных соединений — аденозинтрифосфата (АТФ) и креатинфосфата (КФ); процессов распада и синтеза углеводов, белков и липидов;
окислительных процессов, приводящих в сочетании с гликолизом к ресинтезу АТФ и КФ; происходят синтез и разрушение медиаторов, например ацетилхолина и норадреналина; усиление синтеза РНК и белков.
МАКСИМАЛЬНЫЙ РИТМ ИМПУЛЬСАЦИИ
В естественных условиях существования организма по нервным волокнам проходят не одиночные потенциалы действия, а серии импульсов, следующих друг за другом с различными интервалами. В двигательных нервных волокнах при произвольных движениях частота импульсацци обычно не превышает 50 в секунду, т. е. межимпульсный интервал составляет около. 200 мс. При таком большом интервале все восстановительные процессы, развивающиеся после окончания потенциала действия (реактивация натриевых каналов, восстановление исходной натриевой проводимости, «откачка» из цитопл.азмы ионов Na+ и возвращение внутрь волокна ионов К'+ и т. д.), успевают полностью закончиться. Однако в чувствительных нервных волокнах (например, в слуховом или зрительном нерве) при сильном раздражении в начальный момент частота разряда может достигать 1000 и более импульсов в секунду при длительности абсолютной рефракторной фазы 0,5—0,7 мс. Подобные высокочастотные разряды импульсов появляются при возбуждении и в некоторых нервных клетках, например в клетках Реншоу спинного мозга.
Н. Е. Введенский первый обратил внимание на разную способность возбудимых образований воспроизводить высокие ритмы раздражении. Максимальное число потенциалов действия («максимальный ритм»), которое, способно возбудимое образование генерировать в 1 с в соответствии и с ритмом раздражения, Н. Е. Введенский предложил в качестве показателя «лабильности» ткани. В настоящее время ясно, что максимальный
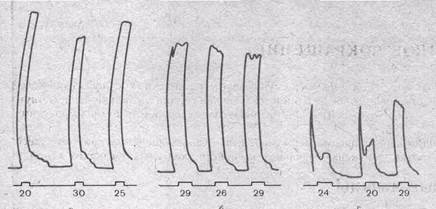
Рис.25. Стадии нарушения проведения через «парабиотический участок» нерва (по Н. Е. Введенскому) .
а — соотношение между силой ритмической стимуляции нормального нерва и высотой тетануса скелетной мышцы. Цифры означают расстояние между катушками индукционного аппарата: чем ближе это расстояние — сильнее раздражение, тем больше частота нервных импульсов и соответственно выше тетанус; бив — то же после смазывания участка нерва 0,5 % раствором кокаина: б— «уравнительная фаза парабиоза»: раздражения разной силы (частоты) вызывают примерно одинаковый эффект; в — «парадоксальная фаза»: сильные (частые) стимулы дают меньший эффект, чем слабые (редкие).
(предельный) ритм импульсации нервных и мышечных волокон определяется скоростями процессов изменений ионной проводимости, лежащих в основе абсолютной и относительной рефрактерности.
Существуют воздействия, замедляющие реактивацию натриевых каналов и потому увеличивающие длительность фазы относительной рефрактерности. К ним относятся, например, местные анестетики. Поэтому участок нервного волокна, подвергнутый воздействию малых концентраций новокаина (или какого-либо другого местного анестетика), утрачивает способность проводить высокочастотные разряды импульсов, тогда как низкочастотные разряды еще продолжают проходить.
При высокочастотной стимуляции происходит либо трансформация ритма (блокируется каждый второй потенциал действия), либо (при очень частой стимуляции) проходит только первый потенциал действия, а остальные оказываются заблокированными. Объясняется это тем, что при частой стимуляции потенциалы действия, приходящие в альтерированный участок, углубляют инактивацию натриевых каналов, вызванную анестетиком.
Сходным образом влияют на процесс реактивации повышение концентрации ионов K.+ в окружающей нервные волокна жидкости и некоторые другие химические агенты.
Н. Е. Введенский (1901) впервые обнаружил нарушение способности нерва проводить высокочастотные разряды импульсов при воздействии на нерв разных химических агентов (рис. 25). Он правильно усмотрел определенное сходство между состоянием, в котором находится нервное волокно при его альтерации химическими агентами, и состоянием рефрактерности, сопровождающей нормальный потенциал действия: и в том и вдругом случае, как это теперь установлено, происходит инактивация натриевых каналов.
Представление о «парабиозе» (так называл Н. Е. Введенский состояние альтерированного участка ткани) как о состоянии «местного неколеблющегося возбуждения» и общей реакции возбудимых образований на повреждающее воздействие и в настоящее время представляет только исторический интерес. Обнаружены агенты, снижающие возбудимость (в результате блокады натриевых каналов), но не влияющие на длительность рефракторных фаз. К числу таких агентов относится, например, специфический блокатор натриевых каналов — тетродотоксин. Имеются также существенные различия в действии агентов, блокирующих проведение, на калиевые каналы и другие транспортные системы мембраны. Таким образом, за внешне сходными изменениями возбудимости и проведения нервных импульсов могут скрываться существенно различные изменения свойств нервного и мышечного волокна.
Глава 3
МЫШЕЧНОЕ СОКРАЩЕНИЕ
У позвоночных животных и человека существует три вида мышц:.поперечно-поло-сатые мышцы скелета, мышцы предсердий и желудочков сердца и гладкие мышцы внутренних органов, сосудов и кожи. Все они различаются строением и физиологическими свойствами.
Свойства мышцы сердца рассмотрены в разделе «Кровообращение», поэтому здесь мы ограничимся только изложением функций и свойств скелетных и гладких мышц.
СКЕЛЕТНЫЕ МЫШЦЫ
ФУНКЦИИ И СВОЙСТВА ПОПЕРЕЧНОПОЛОСАТЫХ МЫШЦ
Померечнополосатые мышцы являются активной частью опорно-двигательного аппарата, включающего, кроме них, кости, связки и сухожилия. В результате сократительной деятельности поперечнополосатых мышц, происходящей под влиянием импульсов, приходящих из ЦНС, возможны: 1) передвижение организма в пространстве; 2) перемещение частей -вела относительно друг друга; 3) поддержание позы. Кроме того, один из результатов мышечного сокращения — выработка тепла.
У человека, как и у всех позвоночных, волокна скелетных мышц обладают тремя важнейшими свойствами: 1) возбудимостью, т. е. способностью отвечать на раздражитель изменениями ионной проницаемости и мембранного потенциала; 2) «проводимостью» — способностью к проведению потенциала действия вдоль всего волокна; 3) сократимостью, т. е. способностью сокращаться или изменять напряжение при возбуждении.
В естественных условиях возбуждение и сокращение мышц вызываются нервными импульсами, поступающими к мышечным волокнам из нервных центров. Чтобы вызвать возбуждение в эксперименте, применяют электрическую стимуляцию.
Непосредственное раздражение самой мышцы называется прямым раздражением;
раздражение двигательного нерва, ведущее к сокращению иннервированной этим нервом мышцы,— непрямым раздражением. Ввиду того что возбудимость мышечной ткани ниже, чем нервной, приложение электродов раздражающего тока непосредственно к мышце еще не обеспечивает прямого раздражения: ток, распространяясь по мышечной ткани, действует в первую очередь на находящиеся в ней окончания двигательных нервов и возбуждает их, что ведет к сокращению мышц. Чтобы получить в эксперименте эффект чисто прямого раздражения, необходимо либо выключить в мышце двигательные нервные окончания ядом кураре, либо приложить стимул через введенный внутрь мышечного волокна микроэлектрод.
Электрическую активность целой мышцы при возбуждении можно зарегистрировать при помощи приложенных к мышце или вколотых в нее электродов и дальнейшего усиления отводимых потенциалов.
Эта методика получила название электромиографии, а регистрируемая с ее помощью кривая — электромиограммы (рис. 26). Последняя представляет собой результат интерференции множества потенциалов действия, асинхронно возникающих в различных мышечных волокнах. Метод электромиографии можно использовать при обследовании человека. Он широко применяется в физиологии спорта и медицине для оценки состояния двигательного аппарата и диагностики ряда заболеваний. Электроды различных типов позволяют отводить внеклеточно потенциалы целой мышцы, отдельных двигательных единиц и даже отдельных волокон. Электромиография позволяет выявлять разнообразные нарушения иннервации мышц и их управления ЦНС.
Для внутриклеточной регистрации мембранных потенциалов отдельных мышечных подокон в эксперименте применяют внутриклеточные микроэлектроды.
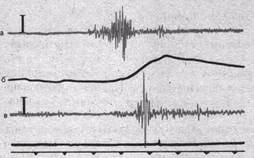 Рис.26. Электромиограмма мышц плеча человека.
а, в — изменения суммарной электрической активности трехглавой и двуглавой (соответственно) мышц плеча; б — изменение локтевого угла. Отметка времени — 10 мс.
Рис.26. Электромиограмма мышц плеча человека.
а, в — изменения суммарной электрической активности трехглавой и двуглавой (соответственно) мышц плеча; б — изменение локтевого угла. Отметка времени — 10 мс.
|
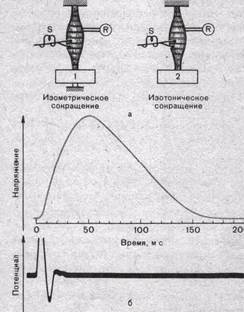 Рис.27. Способы записи изометрического и изотонического сокращений (а) и кривая изометрического сокращения (напряжения) мышцы кошки в ответ на одиночный стимул (б).
S — стимулирующие электроды-, приложенные к двигательному нерву; R—отводящие электроды для записи потенциалов действия мышцы. 1 —тугая пружина с датчиком напряжения; 2 — свободно поднимаемый груз. Под кривой напряжения — запись потенциала действия мышцы.
Рис.27. Способы записи изометрического и изотонического сокращений (а) и кривая изометрического сокращения (напряжения) мышцы кошки в ответ на одиночный стимул (б).
S — стимулирующие электроды-, приложенные к двигательному нерву; R—отводящие электроды для записи потенциалов действия мышцы. 1 —тугая пружина с датчиком напряжения; 2 — свободно поднимаемый груз. Под кривой напряжения — запись потенциала действия мышцы.
|
Регистрация механической активности мышц называется миографией. Для такой регистрации в настоящее время применяют специальные датчики, преобразующие механические изменения (линейные перемещения или напряжение) в колебания электрического тока. После усиления последние могут быть зарегистрированы в виде миограммы (механограммы).
ТИПЫ СОКРАЩЕНИЯ МЫШЦЫ
Потенциал действия, распространяющийся по мышечному волокну, активирует его сократительный аппарат, инициируя акт сокращения. В зависимости от условий, в которых происходит мышечное сокращение, различают два его типа — изотоническое и изометрическое (рис. 27).
Изотоническим называют такое сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным.
Изометрическим называют такое сокращение, при котором мышца укоротиться не может, т. е. когда оба ее конца неподвижно закреплены. В этом случае длина мышечных волокон остается неизменной, а напряжение их по мере развития сократительного процесса возрастает.
Естественные сокращения в организме не бывают чисто изотоническими, так как, даже поднимая постоянный груз, мышца изменяет свое напряжение вследствие реальной нагрузки. Например, при изменении суставного угла руки или ноги меняется плечо рычага, на который действует мышца.
По отношению к целостному организму применяется иная классификация типов сокращения: выделяют изометрическое сокращение, при котором длина мышцы не изменяется; концентрическое, при котором мышца укорачивается, и эксцентрическое, совершаемое в условиях удлинения мышцы (например, медленное опускание груза). В естественных двигательных актах обычно можно наблюдать все три типа сокращения мышц.
Дата добавления: 2016-03-27; просмотров: 4296;