Частоты аллелей. Закон Харди — Вайнберга
В пределах генофонда популяции доля генотипов, содержащих разные аллели одного гена; при соблюдении некоторых условий из поколения в поколение не изменяется. Эти условия описываются основным законом популяционной генетики, сформулированным в 1908 г. английским математиком Дж. Харди и немецким врачом-генетиком Г. Вайнбергом. «В популяции из бесконечно большого числа свободно скрещивающихся особей в отсутствие мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты аллелей сохраняются из поколения в поколение».
Допустим, что в генофонде популяции, удовлетворяющей описанным условиям, некий ген представлен аллелями А1 и А2, обнаруживаемыми с частотой р и q. Так как других аллелей в данном генофонде не встречается, то р+q = 1. При этом q = 1—р.
Соответственно особи данной популяции образуют р гамет с аллелем А1 и q гамет с аллелем А2. Если скрещивания происходят случайным образом, то доля половых клеток, соединяющихся с гаметами А1, равна р, а доля половых клеток, соединяющихся с гаметами A2, — q. Возникающее в результате описанного цикла размножения поколение F1 образовано генотипами AlA1, A1A2, A2A2, количество которых соотносится как (р + q) (р + q) = р2 + 2pq + q2 (рис. 10.2). По достижении половой зрелости особи AlAi и АгА2 образуют по одному типу гамет — A1 или A2 — с частотой, пропорциональной числу организмов указанных генотипов (р и q). Особи A1A2 образуют оба типа гамет с равной частотой 2pq/2.
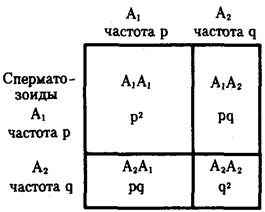
Рис. 10.2. Закономерное распределение генотипов в ряду поколений в зависимости от частоты образования гамет разных типов (закон Харди—Вайнберга)
Таким образом, доля гамет A1 в поколении F1 составит р2 + 2pq/2 = р2 + р(1—р) = p, а доля гамет А2 будет равна q2 + 2pq/2 = q2 + + q(l-q) = q.
Так как частоты гамет с разными аллелями в поколении fi в сравнении с родительским поколением не изменены, поколение F2 будет представлено организмами с генотипами AlA1, A1A2 и А2А2 в том же соотношении р2 + 2pq + q2. Благодаря этому очередной цикл размножения произойдет при наличии р гамет A1 и q гамет А2. Аналогичные расчеты можно провести для локусов с любым числом аллелей. В основе сохранения частот аллелей лежат статистические закономерности случайных событий в больших выборках.
Уравнение Харди—Вайнберга в том виде, в котором оно рассмотрено выше, справедливо для аутосомных генов. Для генов, сцепленных с полом, равновесные частоты генотипов AlA1, A1A2 и А2А2 совпадают с таковыми для аутосомных генов: р2 + 2pq + q2. Для самцов (в случае гетерогаметного пола) в силу их гемизиготности возможны лишь два генотипа A1— или А2 —, которые воспроизводятся с частотой, равной частоте соответствующих аллелей у самок в предшествующем поколении: р и q. Из этого следует, что фенотипы, определяемые рецессивными аллелями сцепленных с хромосомой Х генов, у самцов встречаются чаще, чем у самок.
Так, при частоте аллеля гемофилии, равной 0,0001, это заболевание у мужчин данной популяции наблюдается в 10 000 раз чаще, чем у женщин (1 на 10 тыс. у первых и 1 на 100 млн. у вторых).
Еще одно следствие общего порядка заключается в том, что в случае неравенства частоты аллеля у самцов и самок разность между частотами в следующем поколении уменьшается вдвое, причем меняется знак этой разницы. Обычно требуется несколько поколений для того, чтобы возникло равновесное состояние частот у обоих полов. Указанное состояние для аутосомных генов достигается за одно поколение.
Закон Харди — Вайнберга описывает условия генетической стабильности популяции. Популяцию, генофонд которой не изменяется в ряду поколений, называют менделевской. Генетическая стабильность менделевских популяций ставит их вне процесса эволюции, так как в таких условиях приостанавливается действие естественного отбора. Выделение менделевских популяций имеет чисто теоретическое значение. В природе эти популяции не встречаются. В законе Харди — Вайнберга перечислены условия, закономерно изменяющие генофонды популяций. К указанному результату приводят, например, факторы, ограничивающие свободное скрещивание (панмиксию), такие, как конечная численность организмов в популяции, изоляционные барьеры, препятствующие случайному подбору брачных пар. Генетическая инертность преодолевается также благодаря мутациям, притоку в популяцию или оттоку из нее особей с определенными генотипами, отбору.
Дата добавления: 2016-03-15; просмотров: 748;