ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ (ДРЕЙФ ГЕНОВ)
Мутации и комбинативная изменчивость, периодические колебания численности организмов, изоляция изменяют генофонды популяций случайным образом. Их совместное действие с естественным отбором в процессе видообразования придает биологической изменчивости в целом приспособительный характер. Выполнению отбором упорядочивающей роли препятствуют изменения частот аллелей, зависящие от случайных причин. Таковыми в данном случае являются причины, обусловливающие преимущественное размножение генотипов вне связи с их приспособительной ценностью. Так как динамика частот аллелей в генофондах последовательных поколений носит статистический характер (см. закон Харди — Вайнберга), размах случайных колебаний этого показателя возрастает по мере снижения численности особей в популяции.
Расчеты показывают, что при воспроизведении 5000 потомков родительской популяции с частотой некоего аллеля р = 0,50 колебания концентрации этого аллеля в 99,994% вариантов дочерних популяций в силу случайных причин (в отсутствие отбора по этому аллелю) не выйдут за пределы 0,48—0,52. Если же родительская популяция мала и воспроизводит 50 потомков, то размах случайных колебаний концентрации наблюдаемого аллеля в том же проценте вариантов дочерних популяций составит 0,30—0,70. Случайные, но не обусловленные действием естественного отбора колебания частот аллелей называют генетико-автоматическими процессами или дрейфом генов.
При значительном размахе колебаний в последовательных поколениях создаются условия для потери популяцией некоторых аллелей и закрепления других. В результате происходят гомозиготизация особей и затухание изменчивости. Предположим, что популяция состоит из четырех особей и имеет аллель с частотой р = 0,125. Это означает, что указанный аллель присутствует в генофонде данной популяции в единственном экземпляре у одной из особей, гетерозиготной по соответствующему локусу. Любое случайное стечение обстоятельств, исключающее такую особь из размножения (лесной пожар, выстрел охотника и т.п.), приведет к утрате аллеля. Генофонд дочерней популяции будет его лишен. Вероятность утраты составит 1/2 в случае одного, 1/4 — двух, 1/8 — трех потомков у данной особи. В популяции из 4000 организмов при р = 0,125 минимум 500 особей имеют соответствующий аллель, причем в гомозиготном состоянии. Вероятность исключения всех этих особей из размножения в силу случайных обстоятельств ничтожно мала. Это гарантирует переход аллеля в генофонд следующего поколения и его сохранение.
Дрейф генов обусловливает утрату (р = 0) или закрепление аллелей в гомозиготном состоянии у всех членов популяции (р = 1) вне связи с их приспособительной ценностью. Он играет важную роль в формировании генофондов малочисленных групп организмов, изолированных от остальной части вида.
ВИДООБРАЗОВАНИЕ
Процесс образования видов осуществляется в результате взаимодействия элементарных эволюционных факторов. Видообразование в типичных случаях заключается в разделении первоначально единого вида на два или более новых. Это связано с возникновением межпопуляционных изоляционных барьеров и углублением различий между генофондами популяций под действием естественного отбора вплоть до генетической изоляции. Такой процесс, ведущий к увеличению количества видов, называют дивергентным или истинным видообразованием. Выделяют также филетическое видообразование. Оно заключается в постепенном превращении во времени одного вида в другой. Этот способ наблюдается, если изменения условий захватывают весь ареал. Известны примеры видообразования путем гибридизации. Перечисленные способы видообразования схематически изображены на рис. 11.6.
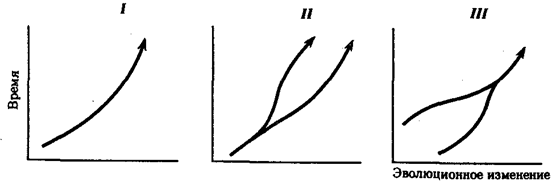
Рис. 11.6. Основные способы видообразования:
I—филетическое, II—дивергентное (истинное), III—путем гибридизации
Существуют аллопатрический и симпатрический пути образования видов. При аллопатрическом видообразовании, называемом также географическим, препятствия к скрещиванию первично обусловлены пространственным разобщением популяций. Генетическая изоляция развивается вторично. Так, некогда в Австралии существовал один вид попугайчиков рода Pachycephala. В условиях засушливого периода единый ареал разделился на западную и восточную зоны. Со временем особи двух популяций приобрели морфофизиологические различия, которые сделали невозможным скрещивание, когда ареал вновь стал общим. Произошло образование из одного предкового вида двух новых.
При симпатрическом видообразовании новый вид образуется внутри ареала исходного вида. С самого начала изоляция является генетической. Такое положение создается в результате полиплоидии вследствие нарушений нормального хода мейоза, при крупных хромосомных перестройках или межвидовой гибридизации. Аллопатрическое видообразование происходит медленно и дает виды, как правило, отличающиеся по морфофизиологическому критерию от вида-родоначальника. Симпатрический путь относительно быстрый и дает виды, близкие к исходному по морфофизиологическим показателям.
Большинство видов, особенно животных, возникают аллопатрическим путем. Симпатрическое видообразование на основе полиплоидии характерно для растений. Так, разные виды пшениц составляют ряд с наборами 14, 28, 42 хромосомы. В клетках дикого хлопчатника 26 хромосом, культурного — 52. Культурная слива возникла путем гибридизации терна с алычой. Примером гибридогенного вида является рябинокизильник, распространенный в лесах центральной Сибири. Симпатрический путь видообразования у паразитов часто связан с освоением популяцией новых хозяев. Анализ генного состава и межхромосомных различий между человеком и человекообразными обезьянами дает повод предположить, что разделение этих двух ветвей могло идти симпатрическим путем.
Дата добавления: 2016-03-15; просмотров: 1011;