Выработка условных реакций и ассоциационизм
Как бы то ни было, изучение феномена памяти в связи с проблемой пластичности и развития нервной системы — это скорее биологический, нежели психологический подход. В экспериментальной психологии первые попытки исследования памяти животных и их способности к научению обычно приписывают Павлову, хотя сам он много позаимствовал у своих предшественников — нейрофизиологов и психологов. По образованию Павлов не был психологом и вопросами научения заинтересовался довольно поздно, уже получив Нобелевскую премию за работы по пищеварению. Всем известно, что он ставил свои эксперименты на собаках, используя звонок в качестве сигнала о приближающемся кормлении. Павлов установил, что выделение слюны, которое обычно происходит, когда перед животным ставят пишу, можно вызвать и с помощью звукового сигнала. Собаки улавливают связь между звуком и последующей дачей корма, что приводит к изменению их поведения. Физиология Павлова была откровенно механистической, и он интерпретировал получаемые результаты весьма прямолинейно. Усадите кого-нибудь с ногой, закинутой на ногу, и слегка ударьте молоточком под коленную чашечку. Результатом будет быстрое подбрасывание ноги — движение вполне бессознательное и неконтролируемое. Нейрофизиологи XIX века, изучавшие жестко детерминированную цепь событий от сенсорного сигнала через нейроны головного или спинного мозга до двигательной реакции, назвали это явление рефлексом. Теория рефлексов для объяснения моторных функций нервной системы уже прочно утвердилась в нейрофизиологии к тому времени, когда Павлов приступил к своим опытам. Поэтому его интересовала реакция собак (слюноотделение на вкус и запах пищи) как фиксированный рефлекс нервной системы, встроенный в систему нейронных связей. Поскольку такой рефлекс проявляется «естественно», без вмешательства экспериментатора, Павлов назвал его безусловным рефлексом. При многократном сочетании кормления со звуком звонка (условным раздражителем) формировался новый рефлекс или нервный путь, который связывал восприятие звукового сигнала и реакцию слюноотделения. Этот рефлекс получил название условного.
Таким образом, существо павловского метода заключалось в сопряжении врожденной реакции с раздражителем, ранее с нею не связанным. В последующие тридцать лет Павлов, а затем и целые поколения его учеников продолжали исследовать природу новой связи 1)*. Они, в частности, выявили важнейшую роль временных отношений: если звонок раздавался после начала кормления или слишком задолго до него, собаки были не в состоянии уловить связь между двумя раздражителями. С другой стороны, удавалось формировать цепочки связей. Если, например, собака усвоила связь между звонками и кормлением, а перед звуковым сигналом включали еще световой, то в конце концов слюноотделение начиналось в ответ на один лишь световой стимул. Такого рода исследования позволили Павлову создать узко-механистическую общую теорию обучения на основе последовательностей («цепочек») условных рефлексов. Пытаясь объяснить более сложные познавательные процессы у человека («высшую нервную деятельность»), он развил в качестве продолжения своей рефлексологии идею о том, что помимо простых условных рефлексов, свойственных животным, у человека имеется «вторая сигнальная система» более высокого порядка. Экспериментальный метод и теоретические формулировки Павлова были с энтузиазмом восприняты большевистской властью. В двадцатые годы политические лидеры и философы коммунистической ориентации испытывали безграничный восторг перед наукой. Материалистический подход Павлова к проблемам психологии привлекал их возможностью исключить всякое участие «души» в мозговой деятельности, а присуждение ученому Нобелевской премии (1904 г.) расценивалось как факт международного признания русской и в дальнейшем советской науки. Ленин лично санкционировал строительство нового большого института, чтобы Павлов мог продолжать работу даже в тяжелейшее время гражданской войны и экономического кризиса в первые годы становления режима, хотя он до самой смерти в 1936 году отказывался вступить в партию или даже просто публично поддержать ее социальную и политическую программу.
*1) Во время моего последнего приезда в получивший свое прежнее название Санкт-Петербург я посетил институт, который теперь носит имя Павлова, и видел его превращенную в музей лабораторию. К немалому своему удивлению я обнаружил по соседству точно такую же действующую лабораторию, где до последних деталей воспроизводятся опыты Павлова с той лишь разницей, что кормление животных и регистрация слюноотделения производятся теперь с помощью компьютерных программ. Увы, это не демонстрационное воспроизведение классических экспериментов; это пример закамуфлированного под науку идолопоклонства.
Вопреки физиологической подготовке Павлова и его подчеркнутому материализму и даже узкому редукционизму в анализе поведения, он проявлял удивительное безразличие к изучению истинно нейрофизиологических процессов, которые могли бы иметь отношение к формированию рефлексов. Он довольствовался изучением феноменологии условных связей и построением абстрактных схем мозговых функций, возможно, лежащих в их основе. В десятилетия власти Сталина и его последователей изучение мозга и поведения в Советском Союзе было загнано в рамки ортодоксальной павловской теории, хотя появлялись новые поколения ученых, которые, отдавая дань бесспорным достижениям Павлова, стремились более свободно развивать теоретические исследования.
Среди наиболее крупных из этих ученых были Выготский, Лурия (чью книгу о Шершевском я уже упоминал), Анохин, С.Л. Рубинштейн и Бериташвили. Выготский умер молодым, а его весьма перспективный социально ориентированный подход к психологии был забыт в мрачные годы сталинизма (в конце тридцатых годов) и обрел новую жизнь только в шестидесятых. Остальные много лет работали, рискуя собственной безопасностью и преодолевая многочисленные трудности, и прожили достаточно долго, чтобы основать собственные научные школы и институты, которые и до сих пор занимают ведущее положение в нейрофизиологии в бывших республиках Советского Союза [5]. При всем скепсисе в отношении отдельных сторон павловского подхода я не могу не отметить важные результаты, полученные в это время советскими учеными в области нейропсихологии, хотя их зачастую игнорируют на Западе по причинам, изложенным в главе 12.
Начиная с двадцатых годов нынешнего столетия павловские условные рефлексы стали обычным предметом изучения и для западных психологов. Однако вскоре это направление сомкнулось с не менее сухим ортодоксальным учением одной из психологических школ США, представители которого называли себя бихевиористами.
Манифест бихевиоризма был провозглашен в 1913 году Джоном Б. Уотсоном. До того времени западная психология мало использовала экспериментальный подход; психологи больше занимались изысканиями в традициях Декарта и пытались понять феномен разума и сознания путем самонаблюдения (интроспекции). В противовес этому бихевиористы стремились полностью исключить из психологии интроспективный метод и само понятие сознания. Они интересовались только внешними, наблюдаемыми аспектами поведения; согласно бихевиористской доктрине, любые предположения о внутренних психических или мозговых процессах, не поддающихся наблюдению, ненаучны и могут быть только предметом веры.
Уотсон и его последователь Б. Ф. Скиннер утверждали, что новорожденный ребенок или детеныш — лишь физиологически полноценный организм, но с поведенческой точки зрения это tabula rasa — «чистая доска», на которой последующий опыт будет оставлять следы, определяющие все дальнейшее поведение. Они полностью сводили поведение к последовательности стимулов и реакций. Организмы на опыте узнают, за какими стимулами следуют приятные раздражители (вознаграждение, или положительное подкрепление), а за какими — неприятные (наказание, или отрицательное подкрепление), и у них вырабатываются соответственные реакции. Формальные термины положительное и отрицательное подкрепление — это элементы сознательной попытки бихевиоризма утвердить себя как абстрактную науку, язык которой не должен ассоциироваться с повседневным опытом. Отсюда следовало, что объекты эксперимента не только не мыслят, но и не «ведут себя», не действуют; вместо того они «выдают элементы поведения», или операнты. Идеальная наука, к которой стремились эти психологи, больше походила на физику, чем на биологию, и служила наглядным примером необыкновенной привлекательности такой весьма нетипичной науки для введенных в заблуждение завидующих представителей других наук.
Олдос Хаксли в своей антиутопии «Этот прекрасный новый мир» пародировал теорию Уотсона и Скиннера, описав выращенных в пробирке методами генной инженерии младенцев, специально сформированных для того или иного места в жизни. В романе Хаксли дети, обреченные стать чернорабочими, получали электрический удар, если пытались прикоснуться к цветам или книгам, что вырабатывало у них глубокое отвращение к чтению и к красотам природы.
Уотсон вскоре сменил карьеру ученого-теоретика на более прибыльное занятие рекламой, и Скиннеру, оставшемуся в Гарварде, пришлось нести дальше факел бихевиоризма, что он и делал на протяжении своей долгой жизни психолога-экспериментатора, советника по вопросам просвещения, философа и романиста, пока не скончался в 1990 году. Выбранный Скиннером путь привел к разработке теории, а потом и методов, которые вышли далеко за рамки павловских. При выработке павловских, или классических, условных рефлексов («классическом обусловливании») подопытные животные не действовали по собственной инициативе, а только реагировали на безусловные и условные раздражители, которые их обучали связывать между собой. Скиннер же помещал животных в такую ситуацию, где от них требовалось активное поведение, т. е. определенное воздействие на окружающую среду. При стандартной форме обучения, которую применяли бихевиористы, животное, например голодную крысу, помещают в ящик с педалью внутри. Когда крыса наступает на педаль — вначале случайно, — ей выдается порция корма, и вскоре она обучается делать это преднамеренно, чтобы получать пищу. Потом поведение животного можно «сформировать» (я опять прибегаю к терминологии Скиннера со всеми ее идеологическими полутонами) таким образом, что оно станет нажимать только на одну из нескольких педалей, нажимать ее строго определенное число раз или, скажем, только после включения света. По аналогии с павловским (классическим) обусловливанием и для отличия от него такое обучение называют инструментальным или оперантным обусловливанием. Очевидно, что здесь возможно множество вариантов: животное можно научить не только производить определенные действия, чтобы получить пищевое вознаграждение, но также выполнять сходные или совсем иные действия, чтобы избежать наказания, скажем, в форме легкого электрического удара (отрицательного подкрепления).
Подобно тому как классический метод стал ортодоксальным в Советском Союзе, поколения студентов-психологов в США и Великобритании (в значительно меньшей степени в других европейских странах) знакомились со своей наукой, наблюдая крыс в скиннеровских ящиках. Последователи Скиннера затратили тысячи человеко-лет, варьируя до мельчайших деталей условия обучения в таких опытах, сравнивая способности крыс, которые для получения корма должны были нажимать на рычаг не один, а несколько раз или повторно на протяжении заданного числа секунд, определяя, как долго животное продолжает нажимать на рычаг, если пища не появляется (затухание реакции). Увлекшись формализацией этих эффектов, сопоставляя разные схемы подкрепления, журналы бихевиористского толка заполняли свои страницы тривиальной феноменологией и перечнями якобы объективных фактов, характеризующих поведение крыс в ящиках; эти факты классифицировались так же тщательно, как марки фанатика-коллекционера, но были столь же далеки, как и при классическом подходе, от биологии и от науки о поведении в более широком контексте. Крысы Скиннера были пустыми черными ящиками, они воспринимали внешние сигналы и реагировали на них как жесткие схемы в компьютере. Как и Павлов, Скиннер был материалистом, стремившимся исключить сознание из своих психологических уравнений, и его материализм, как и у Павлова, был механистичным и редукционистским. Павлов хотя бы довольствовался самой наукой, оставляя ее приложение к проблемам социологии своим последователям, тогда как Скиннер не знал таких ограничений: по-видимому, он даже не замечал уместности пародии Хаксли и не уставал доказывать возможность искусственной психологической организации общества на основе своих теорий поведения и его формирования1.
Скиннер даже утверждал, будто дети обучаются говорить потому, что получают от старших поощрение за правильно произнесенные слова и наказание за ошибки. Эта теория была разгромлена лингвистом Ноамом Хомским в его знаменитой полемике со Скиннером. По мнению Хомского, язык и даже грамматика запрограммированы в «глубинной структуре» мозга, т. е. представляют один из аспектов того, что я называю специфичностью мозга. Подобно тому как в процессе развития глаз образует с мозгом связи, необходимые для упорядоченного зрения, так и речевые центры мозга, по Хомскому, формируют такие связи, какие нужны для создания упорядоченной грамматики. Следовательно, ребенок на период развития запрограммирован таким образом, что обучается говорить грамматически правильно, хотя сам язык, в котором действуют грамматические правила, формируется опытом — той языковой средой, в которой растет ребенок. Я не уверен, что Хомский именно так разъяснил бы свою мысль, но для меня она служит очень удачным примером соотношения специфичности и пластичности в ходе развития [7]. Теория Хомского, однако, недавно подверглась критике с позиций эволюционизма. См. М. Donald, The Origins of the Modern Mind, Harvard, 1991.
Скиннер еще больше, чем Павлов, выхолащивал из живых организмов биологическую сущность, превращая их в артефакты, роботы для психологических манипуляций и тренировок. Правда, «формируя» своих животных, Скиннер обнаружил, что иногда они вдруг ведут себя неожиданным образом. Изредка у них создаются ассоциации, не предусмотренные обучающим: например, животное может подходить к рычагу в определенном направлении или делать какое-то особое движение, прежде чем нажать на него. Скиннер считал такое поведение «предрассудком», результатом случайно возникшей побочной ассоциации, которая привела к ошибочному обобщению. Однако трудно было отрицать, что животные могут что-то узнавать об окружающем мире путем его исследования без специфической связи между особой формой поведения и соответствующим подкреплением — просто благодаря вниманию к происходящему вокруг него. Это можно было бы назвать случайным научением.
Столкновение с фактами реальной жизни животных стало источником чуть ли не замешательства в рядах бихевиористов (один психолог написал даже статью, которую — видимо, не вполне чувствуя иронию, — озаглавил «Недостойное поведение животных»). Так, например, крысы легко обучаются ориентироваться в лабиринте и различать запахи. Голуби, напротив, неважно различают запахи, а лабиринт для них построить трудно, хотя в принципе возможно. В то же время их легко обучить различению цветов и форм и с помощью пищевого подкрепления заставить клевать, например, световые пятнышки красного, а не зеленого цвета. Многие птицы, в том числе почтовые голуби, хотя и не могут ориентироваться в лабиринте, обладают феноменальной способностью строить внутренние карты окружающего мира, находить дорогу к пунктам, удаленным на тысячи миль, или запоминать места, в которых они гнездились или оставили запас корйа. Такие биологические ограничения на способность обучаться, несомненно, связаны с эволюционной историей тех же крыс и голубей, протекавшей в природных условиях. Крысы — ночные животные, они живут в подземных ходах и ориентируются по звукам, запахам и тактильным ощущениям в значительно большей степени, чем с помощью зрения. Птицы же исследуют мир, пользуясь глазами и клювом и не прибегая к обонянию, поэтому некоторые виды ассоциаций создаются у них гораздо легче, чем другие.
Мне потребовалось известное время, чтобы усвоить эту очевидную истину, а когда я узнал, как легко научить цыпленка не клевать повторно горькую бусину, я решил попытаться выработать ассоциацию между видом бусины и каким-то «посторонним» неприятным воздействием. Я задумал эксперимент, в котором всякий раз, когда цыпленок клевал сухую безвкусную бусину, через его ноги проходил слабый электрический ток. Но сколько бы раз клевание бусины ни сочеталось с ощущением от тока, цыпленок продолжал клевать — пожалуй, даже энергичнее, чем раньше, а иногда и несколько агрессивно. В естественных условиях цыплятам вряд ли приходится устанавливать связь между клеванием и болью в ноге, тогда как связь между клеванием и неприятным вкусом может встречаться довольно часто.
Синапсы Хебба
Тем не менее ключевое положение теории Павлова/Скиннера об ассоциативном механизме научения (т. е. о главной роли регулярного сочетания двух тесно сближенных во времени событий, например звонка или нажатия на рычаг с получением корма) было положено в основу концепции научения и несколько десятилетий занимало ведущее место в англо-американских, советских и восточноевропейских традициях (в Западной Европе, где процветали иные научные школы, его роль была гораздо скромнее.) Больше того, понятие ассоциации, вначале чисто психологическое, под пером Доналда Хебба стало уже и нейробиологическим, а книга этого автора «Организация поведения», вышедшая в 1949 году, содержала четкое обоснование клеточного, нейронного варианта ассоциационизма.
Аргументация Хебба сводилась к следующему. Рассмотрим классический павловский рефлекс при обучении собак выделять слюну на звонок. Предположим, что зрительные и обонятельные рецепторы получают сигналы о виде и запахе пищи и передают их дальше по зрительным и обонятельным путям. Эти сигналы через синаптические соединения доходят наконец до специфических нейронов коры. Сигналы от этих нейронов в свою очередь достигают слюнных желез и вызывают слюноотделение. Это, в сущности, такая же рефлекторная дуга, как в случае коленного рефлекса, хотя она значительно сложнее. Но Хебб предположил, что на эти специфические нейроны поступают сигналы не только от зрительной системы, но и от других систем, например от слуховой, с которой те же нейроны образуют слабые, даже нефункционирующие синапсы. Для активации нейронов коры одних только слуховых сигналов может быть недостаточно. Теперь представим себе, что случится, если непосредственно перед кормлением прозвучит звонок. Звуковой и зрительный сигналы поступят на нейроны коры более или менее одновременно. Для активации этих нейронов достаточно зрительного сигнала, но, по мысли Хебба, при частом сочетании обоих сигналов нейроны активируются в такой степени, что у них начинают функционировать и синапсы от слуховой системы. В конце концов нейрон реагирует уже и на один только слуховой сигнал, что вызывает слюноотделение.
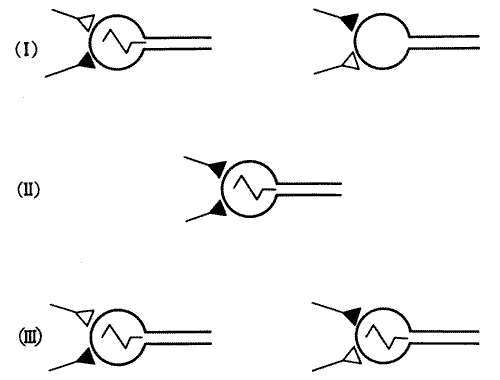
Рис. 6.1. Синапсы Хебба. Согласно Хеббу, следы памяти могут формироваться путем модификации синапсов. На этой схеме показано, как два нейрона образуют синапсы на третьем. Сначала (I), когда активен нижний (темный) синапс, происходит возбуждение третьего нейрона, которое не может быть вызвано через слишком слабый верхний (светлый) синапс. Однако при одновременном воздействии обоих синапсов (II) третий нейрон тоже возбуждается и в нем происходит ряд последовательных биохимических процессов, которые усиливают прежде слабый верхний синапс. В результате (III) верхний синапс приобретает способность сам по себе вызывать реакцию третьего нейрона.
Такое усиление функции синапса могло бы быть результатом биохимической модификации его структуры, изменения электрического потенциала или простым следствием роста. (Разумеется, всегда следует помнить, что, хотя условно говорят об «одном» нейроне, «одном» синапсе и т. д., на самом деле в этот процесс, вероятно, вовлечены сотни нейронов и несколько сотен тысяч синапсов, поскольку число их на один нейрон может достигать тысяч. В обучении даже простой задаче могут участвовать целые клеточные популяции.)
Вот собственные слова Хебба (и его же курсив):
Предположим, что постоянная или повторяющаяся реверберация возбуждения (его «отзвук») вызывает в клетках долговременные изменения, которые усиливают стабильность такого процесса. Это предположение можно точно сформулировать следующим образом: «Если аксон клетки А расположен достаточно близко, чтобы возбудить клетку Б, и постоянно или многократно участвует в активации последней, то в одной или обеих клетках происходят процессы роста или обменные изменения, в результате которых усиливается активирующее действие клетки А на клетку Б».
Согласно наиболее очевидному и, я полагаю, самому правдоподобному предположению о способе усиления возбуждающего действия одной клетки на другую, синаптические соединения разрастаются и увеличивают площадь контакта между пресинаптическим аксоном и постсинаптической клеткой. Разумеется, прямых доказательств правильности такого предположения нет. Однако по целому ряду соображений гипотеза о росте синапсов выглядит вполне приемлемой [8].
Весь этот процесс схематически представлен на рис. 6.1. Мысль Хебба была не нова (я думаю, ни одну научную идею нельзя считать совершенно новой). Предположения такого рода можно найти в работах великого испанского нейроанатома Рамон-и-Кахала, относящихся к началу нынешнего века. Наиболее дотошные молодые специалисты, интересующиеся вопросами приоритета ссылаются даже на статью итальянца Эудженио Танци, опубликованную в 1893 году [9] (другой вопрос, кто из цитирующих эту работу читал ее). Танци был истинным провидцем; в те времена, когда многие еще не признавали нейроны обособленными клетками мозга, он рассуждал следующим образом: Вероятно, каждое отображение чего-либо в мозгу сразу же вызывает функциональную гипертрофию протоплазматических выростов и аксонов; молекулярные вибрации усиливаются и распространяются, изменяя форму дендритов; таким образом, при благоприятных обстоятельствах возникают, а потом и сохраняются новые выросты и ответвления... Каждый последующий эпизод одного и того же акта сознания в форме воспоминания, фантазии или идентичного повторения усиливает мнемонический потенциал, поскольку ведет к формированию новых впечатлений, которые заменяют или дополняют существовавшие ранее... При этом на смену первичным кратковременным связям между нейронами в конце концов приходят стабильные функциональные объединения клеток... Этот прогрессирующий процесс функциональной гипертрофии приводит к более или менее стойкому увеличению разветвленных отростков, соединяющих нервные элементы [9].
Идеи Хебба были высказаны в то время, когда стала уже появляться возможность их экспериментальной проверки, поэтому сразу же после выхода его книги «синапсы Хебба» и «хеббовы правила ассоциации» послужили, как отмечалось в главе 4, исходным материалом для построения моделей и теорий памяти. Большинство из нас, ведущих экспериментальные исследования памяти, полагает, что механизмы вроде постулированных Хеббом действительно должны существовать; трудность состоит в том, чтобы подтвердить это и выяснить, как именно происходят биохимические, физиологические и структурные изменения.
В модели Хебба очень привлекает, то что она (как считали его современники и до сих пор считают многие исследователи) предлагает способ прямого «перевода» поведенческого аспекта ассоциации (классического обусловливания) на язык нервных механизмов. Она объясняет видовые различия в способностях к научению, так как, согласно ее центральному постулату, для усиления функции синапсов необходимы по крайней мере потенциальные синаптические связи (в приведенном выше примере из павловских опытов — связи между нейронами, получающими слуховые сигналы, и нейронами, запускающими слюноотделение). Это существенно ограничивает возможности формирования любой специфической ассоциации. Не исключено, что у моих цыплят не было таких потенциальных связей между нервными клетками, реагирующими на электрическое раздражение ноги, и клетками, «запрещающими» клевать тот или иной предмет.
Назад к организму
При всей популярности ассоциаций среди психологов-теоретиков, разработчиков моделей и нейробиологов-экспериментаторов они не могут быть единственным механизмом памяти, на что уже в двадцатых годах указывали многие критики теорий Павлова и Скиннера. Например, по теории Скиннера крысы должны осваивать лабиринт, запоминая последовательные повороты (сначала налево, потом направо и т. д.) как звенья в цепи стимулов и реакций. Однако перестраивая лабиринт или изменяя в нем ориентиры, нетрудно было показать, что стратегия животных не столь примитивна. После нескольких тренировок в их мозгу формировалось нечто вроде цельного общего представления об устройстве лабиринта — если угодно, его карта. Поэтому в какую бы его часть ни помещали животное, оно быстро догадывалось, где должна находиться приманка, и выбирало самый короткий путь к ней, не испытывая большого замешательства в связи с перестройкой лабиринта. В процессе обучения животные вырабатывают определенные стратегии и формируют концепции. Для понимания таких механизмов недостаточно изучать лишь последовательность ассоциаций «стимул—реакция» и применять положительное или отрицательное подкрепление.
В двадцатые и тридцатые годы этот более широкий взгляд на механизмы поведения животных послужил основой для создания школы гештаяьтпсихологии, получившей особое развитие в Германии и Австрии. Если Скиннер и Павлов были редукционистами, то гештальтисты исходили из идей холизма. Прототипом их экспериментов по обучению животных стали опыты, проводившиеся Вольфгангом Кёлером на шимпанзе. Обезьяну помещали в клетку, откуда она видела банан, но не могла дотянуться до него; в клетке имелись две палки, каждая из которых была слишком короткой, чтобы с ее помощью достать банан. Обезьяна должна была догадаться, что для получения корма следует соединить две палки в одну. Кёлер и гештальтисты полагали, что решение этой задачи свидетельствует о творческом, концептуальном мышлении шимпанзе, а не просто о способности объединять длинные цепи стимулов и реакций [10]. Это альтернативное направление в сочетании с идеями генетической эпистемологии, которые в те же годы развивал швейцарский психолог Жан Пиаже, составило серьезную конкуренцию ассоциационизму, особенно в Западной Европе. Конкуренция еще больше усилилась в связи с повышенным интересом английских и других европейских исследователей к изучению поведения животных в условиях более естественных, чем те, которые существуют в искусственно ограниченном пространстве лабораторных клеток и лабиринтов. Это направление, получившее название этологии, вскоре начало заново открывать все разнообразие поведения животных, которому не было места в экспериментах Скиннера и Павлова и которое редукционизм пытался изгнать из своих построений [11].
Именно это различие дало повод (кажется, Бертрану Расселу) заметить, что разница между немецкими и американскими крысами состоит в том, что в Америке животные снуют, обучаясь методом проб и ошибок, а в Германии сначала сидят и обдумывают свои действия. Между прочим, гештальтисты проявляли еще меньше интереса, чем другие школы добиологической психологии, к физиологическим процессам, происходящим в мозгу у их мыслящих обезьян.
Вот почему я берусь утверждать, что нейробиологи, занимающиеся памятью и научением, получили в наследство от того периода экспериментальной психологии (если не считать идей Хебба) не теоретические построения, не накопленные с таким трудом феноменологические данные и не разработанные до мельчайших деталей процедуры обусловливания и подкрепления. Если бы александрийский пожар уничтожил тысячи метров библиотечных полок с хранившимся на них архивом бихевиористских и павловских журналов за двадцатые — шестидесятые годы, то, я убежден, мы утратили бы в основном материалы, представляющие только исторический интерес. При всем внимании Павлова и Скиннера к теории их влияние еще сказывается лишь на методических подходах: еще несколько лет назад почти во всех экспериментально-психологических лабораториях использовали ящики Скиннера, а также некоторые модели классического обусловливания (но теперь и они выходят из употребления, уступая место опытам с задачами, требующими более четкого проявления познавательных способностей и создания «внутренних карт», о чем речь пойдет позже).
Если бы меня попросили указать наиболее важный поворотный момент в истории американских исследований по проблеме научения, я без колебания назвал бы эксперименты, проводившиеся Джоном Гарсиа в конце пятидесятых — начале шестидесятых годов. Гарсиа заинтересовался «боязнью приманки» — феноменом, хорошо известным каждому, кто когда-нибудь пытался избавиться от нашествия мышей с помощью отравленных приманок или ловушек. Если животное найдет отравленный корм и съест его, а потом будет несколько часов, чувствовать себя больным, то в дальнейшем оно не станет брать приманку. Если данный вид пищи знаком животному, но встретился ему в необычном месте, оно будет избегать кормиться в этом месте. Если же необычный (по вкусу или виду) корм был найден в знакомом месте, животное вообще перестает брать такой корм, где бы он ни оказался. Гарсиа смоделировал боязнь приманки экспериментально. Он использовал безвредный корм, но либо предлагал его в непривычном месте, либо придавал ему необычную окраску, а затем вызывал у животных болезненное состояние, вводя в желудок небольшое количество хлористого лития; это вызывало такие же ощущения, как при легком отравлении приманкой, и они сохранялись в течение трех часов. После нормализации состояния эти животные избегали брать корм необычного цвета или в новом месте; у них явно создавалась ассоциация между поеданием такой пищи и последующими болезненными ощущениями. (Сходное явление известно и у человека: при проведении химиотерапии в условиях стационара у больных нередко развивается отвращение к больничной пище [Первые исследования Гарсиа финансировало военно-морское ведомство США, интересовавшееся тем, как влияет на аппетит облучение (например, после ядерных взрывов); его влияние сходно с действием химио- или радиотерапии.]) [12].
Описанное поведение настолько знакомо всем, кто работал с животными (или с людьми), что можно было бы удивляться, почему оно вызвало такую растерянность у психологов — приверженцев ассоциационизма. Однако объясняется это довольно просто. Согласно обычным правилам классического и оперантного обусловливания, для выработки ассоциаций необходима достаточная близость во времени между условным и безусловным раздражителями. Условный стимул должен опережать появление безусловного не более чем на четверть секунды; очень коротким должен быть и интервал между действием животного и подкреплением при оперантном обусловливании, иначе ассоциация не установится. Однако при условнорефлекторном избегании пищи (так стали называть феномен Гарсиа) недомогание, т. е. отрицательное подкрепление, может появляться через несколько часов после кормежки. Ассоциационисты, как и следовало ожидать, всеми силами старались спасти свою теорию. Может быть, в то время, когда животные чувствовали себя плохо, формировалась ассоциация с неким остаточным ощущением вкуса пищи, находившейся в желудке? Однако этот аргумент не выдерживает критики, поскольку реакция избегания вырабатывается и по отношению к необычному месту, где была съедена обычная по вкусу пища.
Пару лет назад я невольно оказался вовлеченным в эти споры, когда разрабатывал вариант теста Гарсиа для цыплят. Я предлагал им смоченную водой зеленую бусину, которую, как я и ожидал, они энергично клевали. Через полчаса я вводил им хлористый литий, они слегка заболевали, потом выздоравливали, и спустя три часа им предъявлялась та же бусина. Разумеется, они не трогали ее, они уже знали, что «зеленая мокрая бусина» нехороша. Тогда я видоизменил план эксперимента и вместо влажной давал цыплятам сухую зеленую бусину. Они клевали ее, потом, как и раньше, заболевали и в последующем отвергали зеленую бусину, хотя продолжали клевать красную и хромированную. Очевидно, что в этом случае не могло идти речи о каком-то остаточном вкусе, который «ассоциировался» с болезненным состоянием. Возможно было только одно объяснение: у цыплят, впервые клевавших новую бусину, формировалось некое «представление» о зеленой бусине — «модель», которая удерживалась в мозгу по меньшей мере полчаса и в это время не сочеталась ни с приятным, ни с неприятным ощущением, была просто нейтральной. В последующем у них создавалась ассоциация (теоретически незаконная!) между этим возникшим ранее представлением о бусине и ощущением дискомфорта, что и заставляло их отвергать после этого зеленую бусину [13].
Хотя я не собирался участвовать в упомянутом споре об ассоциационизме, я не мог не видеть, размышляя о возможном значении моих экспериментов, что при всей ясности их результата он не укладывался ни в одну из простых ассоциативных концепций, основанных на условнорефлекторной теории Павлова и Скиннера с их особым упором на очень малый разрыв во времени между стимулом и реакцией. Правда, ассоциационисты могли выдвинуть тот аргумент (некоторые действительно прибегали к нему, когда я обсуждал с ними свои опыты), что запоминание связи между видом бусины и реакцией на нее объяснялось ассоциацией не между клеванием и вкусом, а между клеванием и зеленым цветом или, может быть, между формой и зеленым цветом бусины, и т. п.
Очевидно, можно придумать множество других подобных пар, и будет очень трудно опровергнуть такие гипотезы. Но все они не имеют отношения к сути ассоциативной теории. Если следовать представлениям Павлова и Скиннера, связываться должны не просто два свойства одного предмета (скажем, зеленая окраска с округлой формой), а либо свойство предмета с подкреплением, таким как приятный вкус или плохое самочувствие, либо действие животного (например, клевок) с его немедленным следствием (например, неприятным вкусом). Сказать, что цыплята связывают нейтральный акт — клевок — с каким-то предметом, одновременно зеленым и круглым, без какого-либо положительного или отрицательного подкрепления значит сказать лишь то, что животное, помнящее бусину, способно узнавать отдельные ее особенности, в том числе цвет и форму. В главе 11 я собираюсь показать, как наши собственные эксперименты подтверждают это, и вывести заключения, весьма отличные от тех, которые хотели бы сделать ассоциационисты.
Из всего сказанного можно извлечь трю урока. Во-первых, значение простых экспериментов нередко может оказаться значительно большим, чем ожидалось при их планировании. Во-вторых, как указывали многие философы науки, теории отмирают с трудом. В-третьих, у животных (даже однодневных цыплят) есть разные способы познания мира, поэтому наши теории должны быть достаточно широкими, чтобы охватывать все эти возможности. Нельзя говорить, что теория ассоциаций неверна — она просто не исчерпывает проблему.
Однако вместо того, чтобы дальше обсуждать все повороты и изгибы психологического теоретизирования по вопросам памяти, я хочу вернуться к ее биологическим аспектам. Я рассматриваю память как особую, весьма специализированную форму приспособления к среде, которое свойственно всем живым организмам. Для выяснения механизмов человеческой памяти я предложил изучать процессы научения и вспоминания у животных — кошек, собак, крыс, голубей или цыплят. Все это позвоночные с большим, сложно организованным мозгом. Связано ли наличие памяти с большими размерами мозга, или животные с гораздо проще устроенной нервной системой тоже могут обучаться и вспоминать? Обучаются ли какие-то животные лучше других, и если так, то нет ли в их мозгу некой специализированной структуры или особых биохимических и физиологических механизмов, отсутствующих у видов, менее способных к этому? Ответы на эти вопросы, возможно, прольют свет на механизмы памяти, так как помогут выяснить, какие свойства мозга необходимы для ее формирования и проявления. В связи с этим нам полезно будет рассмотреть эволюцию памяти и формы, которые она может принимать у животных с менее развитой и не столь сложной, как у позвоночных, нервной системой и даже у организмов, вовсе ее лишенных. Этому посвящена следующая глава.
Глава 7
Эволюция памяти
Есть небезобидная склонность, в которой нередко повинны и нейробиологи: часто говорят о некой эволюционной лестнице или шкале сложности, вдоль которой можно разместить все ныне живущие на Земле организмы — от самых "примитивных" до самых «высокоразвитых». Еще более крайняя форма такого мышления предполагает действие в эволюционном процессе какого-то рода направляющей силы, сделавшей человека вершиной прогрессивного развития. В основе подобных идей, которые берут начало в представлениях о месте человечества в природе, сложившихся задолго до Дарвина и появления современной биологии, лежит глубокое непонимание эволюции. Поэтому дальнейшему обсуждению придется предпослать краткую вступительную лекцию. Все современные организмы произошли от примитивных форм жизни, появившихся на Земле довольно скоро после рождения самой планеты, вероятно около четырех миллиардов лет назад. Под эволюцией (это слово буквально переводится как «развертывание» или «развитие») биологи понимают процесс постепенного изменения форм организмов в ряду поколений. «Форма» здесь означает все присущее организму — от биохимических и структурных особенностей до поведения. Изменения формы происходят на самых разных уровнях, от молекулярного до популяционного, и сохраняются благодаря наследственной передаче 1). Когда изменение становится настолько большим, что уже препятствует успешному скрещиванию новой формы со старой, можно говорить о возникновении нового вида.
*1) Я дал лишь самое общее определение эволюции. Мое описание формы включает и то, что генетики называют фенотипом организма (т. е. все, кроме генов), и его генотип (полный набор имеющихся у организма генов). Популяционные генетики обычно исходят из гораздо более строгой редукционистской формулировки, определяя эволюцию, так сказать, с точки зрения гена: они считают самым существенным изменение генных частот, т. е. интересуются прежде всего тем, как часто встречается тот или иной ген в популяции организмов одного вида Я не хотел бы сейчас вдаваться в обсуждение всех этих различий, которое многими будет воспринято как продолжение текущих споров между мною и многими другими, с одной стороны, и такими социобиологами, как Докинз и Уилсон, — с другой. Ричард Левонтин, Лео Камин и я подробно развили эту тему в другой публикации [1].
Самый общепризнанный механизм эволюционного изменения — это современная версия дарвиновского естественного отбора, основанная на следующих простых посылках: а) организмы порождают себе подобных, хотя и с небольшими, главным образом случайными, вариациями; б) все организмы дают больше потомков, чем может дожить до зрелого возраста и в свою очередь приступить к размножению; в) выжившее и размножающееся потомство представлено особями, лучше приспособленными к окружающим условиям, чем погибшие особи; г) эти выжившие особи имеют больше шансов дать лучше приспособленное потомство; таким образом, новые, более удачные формы будут постепенно вытеснять менее приспособленных предшественников.
Живые организмы могут приспосабливаться к существующим условиям, но не могут прогнозировать будущее. Вероятность того, что последующие поколения будут жить в более теплом климате, не может повлиять на отбор в настоящем. Таким образом, эволюционное изменение в популяции организмов способно отражать только те изменения среды, которые уже произошли. Эволюция не обладает даром предвидения. В то же время среда никогда не бывает статичной, она постоянно меняется. Физические силы изменяют климат, вызывают образование гор и опускание суши. Живые организмы и сами влияют на собственную среду обитания, изменяя ее химический состав в результате потребления питательных веществ, дыхания и выделения продуктов жизнедеятельности; они служат факторами географических изменений, разрушая и созидая новые элементы ландшафта (эта довольно-таки очевидная концепция получила почти метафизическое истолкование в работах Джеймса Лавлока и его приверженцев под претенциозным названием гипотезы Геи). Помимо всего прочего, каждый организм составляет часть окружающей среды для других организмов, выступая в качестве полового партнера, родителя, потомка, хищника или добычи. Наконец, организмы не пассивные реципиенты внешних воздействий: они (животные в большей степени, чем растения) могут выбирать для себя условия жизни, перемещаясь из менее благоприятной среды в более благоприятную. Таким образом, в эволюции, как и в процессе индивидуального развития, связь между организмом и средой всегда двухсторонняя: среда непрерывно бросает вызов организму, и тот либо противостоит ей (успешно размножается), либо не выдерживает (вымирает). Организм и среда диалектически взаимосвязаны.
Дарвиновский отбор — это, вероятно, лишь один из возможных механизмов эволюционного изменения живых существ (об относительной роли других процессов продолжается оживленная дискуссия, но она выходит за рамки нашего обсуждения). Сейчас важно подчеркнуть, что все современные формы земной жизни, несомненно, сложились на основе сходных требований, предъявлявшихся отбором на протяжении всего геологического времени. Нельзя поэтому сказать, что один вид «более развит» или «лучше приспособлен», чем другой. Если навозный жук, гриб или человек до сих пор существуют, то это само собой означает, что они сумели сохраниться как виды. Есть много разных способов выживания и продолжения рода, и для каждого организма существуют свои критерии эволюционного «успеха». Только очень предубежденный, рассудочный взгляд на мир живого может подсказать мысль о прямой связи выживания с размерами мозга. Если исходить из общего объема биомассы, окажется, что организмов, не имеющих мозга (или даже вообще центрального нервного образования), несравненно больше, чем животных, обладающих им. Даже среди животных с высокоразвитой нервной системой жуки явно преобладают по численности над млекопитающими 1).
*1) Не могу удержаться, чтобы не процитировать здесь ответ Дж. Холдейна — физиолога и генетика марксистского толка, который в двадцатых — тридцатых годах внес большой вклад в новую синтетическую теорию эволюции. Когда его спросили, какие, по его мнению, качества свойственны Богу, Холдейн сказал: «Бог, должно быть, необычайно любит жуков».
Нельзя говорить о прогрессе или целенаправленности эволюции. Примитивные одноклеточные существа — наши ранние предшественники — не обладали собственным стремлением и не побуждались высшей мистической силой к развитию, которое привело к появлению человека. Правда, существуют строгие ограничения на то, что может и что не может развиваться, связанные с физическими условиями нашей планеты и с химией углерода, составляющей молекулярную основу жизни на Земле. Но в пределах этих ограничений происходили случайные события, изменялись отдельные организмы в определенное время и в определенных местообитаниях, и все это привело в конце концов к современным «высшим» формам жизни. Не было никакого генерального плана. Изучение ископаемых остатков дает представление о том, как складывалась эволюция. Но, как замечает один из последних комментаторов, Стивен Дж. Гоулд [2], если бы можно было вернуть колесо эволюции обратно к моменту зарождения жизни на Земле, а потом заново раскрутить его, результат скорее всего был бы совсем иным: не возник бы, вероятно, ни человек, ни даже мозг. Изучение эволюции — это изучение истории, а не разработка некой математической программы продвижения от простого к сложному и не разгадка внутреннего стремления к эволюционному совершенствованию в духе Тейяра де Шардена 1). В этой исторической последовательности некоторые организмы, например человек, появились совсем недавно, а их предки, судя по ископаемым остаткам, очень быстро претерпели коренные изменения. Другие формы жизни на Земле гораздо больше напоминают своих предков, живших много миллионов лет назад. Создается впечатление, что, однажды найдя способ успешно выживать и размножаться в относительно стабильной среде, они в дальнейшем не нуждались в существенном изменении образа жизни. Поэтому, изучая их биологию и поведение, можно с достаточной долей уверенности заключать о том, как жили их (а значит, и наши) давние предки.
*1) Последняя сноска, касающаяся эволюции. В нескольких фразах последнего абзаца кое-кто, возможно, увидит скрытый намек на дружеский спор, который я вот уже несколько лет веду с моими коллегами по Свободному университету — Брайеном Гудвином и Мэ-Вань Хо, убежденными сторонниками (наряду с Джерри Уэбстером и Питером Сондерсом) такого рационалистического, в противовес историческому, подхода к пониманию биологической формы [3]. Я разделяю мнение большинства биологов, что в этой науке ничто не имеет смысла вне исторического контекста, хотя одновременно соглашаюсь, что любое явление в биологии возможно только в рамках структурных ограничений, налагаемых физическими принципами. Иными словами, я занимаю промежуточную позицию между взглядами Гоулда и Гудвина с компанией. Согласно первому, все решает случай: непредсказуемые события, будь то мутации или массовое вымирание вида вследствие изменившихся условий среды, определяют будущее развитие, которое можно понять только в ретроспективе («война проиграна... и все из-за гвоздя»). Для Гудвина и его единомышленников изучение истории — это рутина («не более чем собирание древностей», по выражению Уэбстера); если достаточно хорошо понять формообразующее действие физических и химических сил, то нетрудно убедиться, что число возможных типов живых организмов крайне ограниченно и появления человека в общем-то можно было ожидать.
Возвращаясь далеко назад, к истокам эволюции, мы видим, что все современные организмы имели общего предка или группу предков. На пути, который привел к появлению человека, находились существа, жившие три (или более) миллиона лет назад и бывшие также предками современных человекообразных обезьян. Еще раньше существовали предки всех современных млекопитающих, а до них — предки всех ныне живущих позвоночных; наконец, мы дойдем до предков всех многоклеточных организмов, и т. д. Эти отношения лучше всего представлять не в виде эволюционной лестницы или дерева, а в виде куста с многочисленными веточками. Строение мозга и поведение предков современных животных можно представить себе, только изучая их живых потомков, поскольку в ископаемых остатках не сохранились мягкие ткани, такие как мозг или нервы. Этот сравнительно-психологический метод позволяет тем не менее прийти к некоторым выводам об эволюции нервной системы, мозга, поведения и, наконец, памяти.
Истоки поведения
Адаптивное поведение как обязательное условие выживания свойственно всем организмам, однако многие процветающие формы жизни прекрасно обходятся без научения и вспоминания, даже без мозга. Возьмем подсолнечник, соцветие которого всегда обращено к источнику света, т. е. энергии. По мере роста подсолнух не учится на опыте, как лучше поворачиваться, и он не перестанет делать это, если после каждого поворота «наказывать» его электрическим ударом. Это поведение (поворачивание соцветия), носит оно адаптивный характер или нет, Дано организму от природы, зафиксировано в его генетическом коде и программе развития. Такое присущее виду поведение (обычно говорят врожденное, но я избегаю пользоваться этим термином, так как он несет дополнительную идеологическую нагрузку) гарантирует адекватный ответ на специфические раздражители без научения методом проб и ошибок, но за счет ограничения диапазона и гибкости реакций. Поведение изменяется не только у отдельной особи, но и в длинном ряду поколений как следствие эволюции. В отличие от этого приобретенные формы поведения вырабатываются на опыте, на практике; их преимущества — большая гибкость, свобода корректировки при изменениях окружающей среды и возможность учитывать наблюдаемые последствия при жизни данного индивидуума. Очень важно, однако, помнить, что приобретенные путем научения реакции не наследуются потомством. Передаются лишь степень пластичности и способность обучаться, которые в свою очередь «заданы» генетическим кодом и программой развития. Вот почему пора забыть наскучившие старые споры о врожденном и приобретенном. Чтобы познать мозг и поведение, нужно отрешиться от этого противопоставления и попытаться вместо того понять диалектическую связь между специфичностью и пластичностью.
Способ существования растений определяется их способностью выживать, используя энергию Солнца для усвоения двуокиси углерода и преобразования ее в нужные питательные вещества, в частности в сахар. Для этого надо лишь оставаться на месте с раскинутыми ветвями, осуществлять фотосинтез и стараться избегнуть поедания, чтобы успеть произвести потомство. За это приходится расплачиваться большой сложностью способов размножения, которые по самой своей сути не очень надежны, так как зависят от других организмов или природных сил (например, ветра), разносящих пыльцу и семена. Жизнь животных не столь проста. Не обладая способностью к фотосинтезу, они вынуждены использовать частично готовую пищу, либо непосредственно поедая растения, либо охотясь на животных, питающихся растениями. Для этого им приходится непрерывно передвигаться, прибегать к специализированным способам поиска корма, спасаться от опасностей и разыскивать партнеров. Таким образом, неспособность к фотосинтезу компенсируется расширением репертуара адаптивного поведения и его большей гибкостью, развитием органов чувств, позволяющих обнаруживать пищу и врагов на расстоянии, и двигательными навыками для перемещения в желаемых направлениях.
Одиночные клетки
Некоторые из описанных форм поведения встречаются даже у простейших одноклеточных организмов. Еще в 80-х годах прошлого столетия в Германии В. Пфейфер провел следующий эксперимент. Он опускал тонкую капиллярную трубочку, заполненную раствором глюкозы, в каплю жидкости, содержавшей бактерии, и наблюдал, как они концентрировались у отверстия капилляра. Бактерии вели себя так, будто «знали», где находится глюкоза, и демонстрировали адаптивную реакцию. Объясняется это довольно просто: глюкоза медленно диффундировала из капилляра в окружающую жидкость, и здесь создавался градиент концентрации сахара. В мембранах бактериальных клеток имеются рецепторные молекулы — белки, строение которых позволяет им узнавать молекулы глюкозы, и крошечные бичевидные жгутики бактерий, работая синхронно, перемещают клетку «вверх» по градиенту. У мутантных форм, лишенных рецепторов, такое поведение отсутствует, что не мешает им использовать глюкозу как источник энергии, если их поместить в раствор.
Более сложные одноклеточные организмы, например парамеции, имеют выросты иного типа, называемые ресничками. Основания ресничек соединены системой тонких нитей, координирующих их биение. В интервалах между эпизодами питания (добычей им служат бактерии) парамеции находятся в непрестанном движении и часто наталкиваются на препятствия, после чего меняют направление движения, заставляя реснички работать на той или другой стороне тела. Таким образом эти животные, перемещаясь подобно игрушечным машинкам, обходят препятствия и достигают участков, богатых пищей. Точно так же они способны уплывать из зон со слишком высокой или низкой температурой или избегать таких раздражающих веществ, как кислоты. Эти реакции поиска и избегания у существ размером не более 0,2 мм, не имеющих нервной системы, есть не что иное, как зачатки более сложных форм активного поведения.
Дата добавления: 2016-03-05; просмотров: 1037;