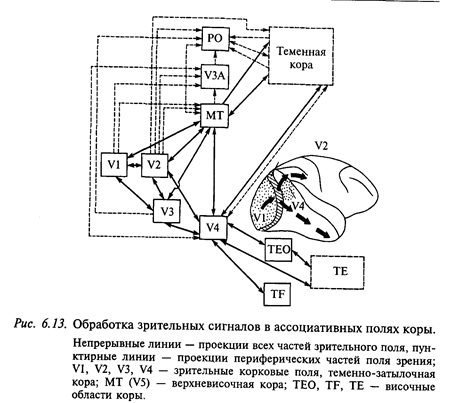
Обработка зрительных стимулов в ассоциативных полях коры.
Исследование мозга низших обезьян показало, что анализ зрительной информации не завершается в полях зрительной коры (17, 18, 19). Было установлено, что от поля 17 начинаются пути (каналы), в которых производится дальнейшая обработка зрительных сигналов. Один путь, идущий в дорсальном направлении к полям теменной ассоциативной коры, участвует в формировании пространственного зрения, другой путь - вентральный (нижневисочная кора) участвует в формировании предметного зрения. В дальнейшем мы будем здесь применять классификацию зрительных корковых полей, принятую в современной литературе. Согласно этой классификации поле 17 (по Бродману) соответствует полю VI (от англ. Visual - зрительный), поле 18- V2, поле 19- V3;
Поля V4 и V5 в этой классификации не имеют соответствия полям карты Бродмана.
Как видно из приведенной схемы (рис. 6.13), зрительная информация поступает к нейронам поля V4 (находится на стыке височной и теменной областей) по нескольким каналам. В поле V4 имеется весьма грубая ретинотопия. Локальное разрушение этого поля у обезьяны нарушает константное восприятие цвета, но не изменяет восприятие формы. Однако при более обширных повреждениях этой зоны у обезьян страдает восприятие как константности цвета, так и формы. Обработка зрительной информации о форме, как предполагают, происходит в нижневисочном поле. Эксперименты с локальным повреждением этой зоны, а также регистрация реакций отдельных нейронов показали, что нейронные сети здесь, по-видимому, участвуют в таких функциях, как зрительное дифференцирование различных объектов во внешней среде. На всем пути от VI к V4 и далее к нижневисочному полю наблюдается устойчивая тенденция к увеличению площади рецептивных полей отдельных нейронов.
Поведенческие эксперименты на обезьянах показали, что разрушение нижневисочного поля приводит к потере способности узнавать зрительные объекты. Предполагают, что это обусловлено потерей способности устанавливать эквивалентность изображения от одного и того же объекта, если он проецируется на разные участки сетчатки. Исследование реакций нейронов нижневисочной области показало, что они лучше реагируют не на простые стимулы (точки, полоски и пр.), а на объекты со сложным контуром. Например, в этой области обнаружены нейроны, которые избирательно активировались при предъявлении в качестве зрительного стимула изображения лица обезьяны. Наиболее часто нейроны такого типа обнаруживаются в передней височной области коры. Биологическое назначение такой важной способности, как узнавание лица, у приматов совершенно очевидно: именно это, по-видимому, лежит в основе социальных функций коммуникации, определения принадлежности к группе и т. д. Наблюдения за эффектами разрушений нижневисочных областей коры приводят исследователей к выводу, что в этом случае не страдают базисные свойства восприятия, такие, как острота зрения и восприятие цвета. Вместе с тем выходят из строя механизмы высшего уровня анализа. Одно из предположений состоит в том, что нарушается выделение объектов какой-то одной категории. Например, страдает различение лиц в категории, которую можно обозначить как «лица обезьян». По мнению некоторых ученых, функция зрительной памяти нижневисочной коры существенно усиливается влияниями со стороны миндалины и гиппокампа.
Роль верхних двухолмий в анализе зрительных стимулов и движении глаз. Ориентация головы с находящимися в ней глазами на зрительные стимулы, которые появляются в поле зрения, играет большую роль в поведении позвоночных животных, включая человека. Особенно сильную ориентировочную реакцию вызывают движущиеся объекты. Одной из ведущих структур, обеспечивающих ориентировочное поведение, является верхнее двухолмие. Верхние слои верхнего двухолмия обезьяны содержат нейроны, реагирующие на появление небольших зрительных стимулов. Подавляющее число нейронов этой области реагируют на движение в любом направлении, и лишь около 10% нейронов верхнего слоя реагируют на движение стимула в одном, предпочтительном направлении (дирекционно селективные нейроны). Эти два класса нейронов ослабляют свой ответ при раздражении стимулами большой площадью. Это свидетельствует о наличии тормозной зоны, окружающей центральную возбудительную зону рецептивного поля. В верхних слоях двухолмия имеется упорядоченная проекция сетчатки (ретинотопия).
При погружении микроэлектрода вертикально поверхности двухолмия местоположение зрительных рецептивных полей не изменяется (колончатая организация), но рецептивные поля нейронов, расположенных более глубоко, как правило, большего размера. Отмечено также, что чем ближе рецептивные поля нейронов к fovea, тем меньше их угловые размеры, а чем дальше от fovea, тем они больше (до 20°).
В нижних слоях серого вещества двухолмия находятся нейроны, которые не реагируют на зрительные стимулы, но имеют так называемые моторные поля, т.е. нейрон максимально активируется при саккаде глаза в определенном направлении. Активация нейронов этого слоя двухолмия всегда опережала на десятки миллисекунд саккады глаза. При этом реакция нейрона была одинаковой независимо от способа вызова движения глаза (предъявление зрительного стимула в определенной части зрительного поля, спонтанном движении глаза в темноте или нистагме, вызванном стимуляцией вестибулярного аппарата). Локальная электрическая стимуляция двухолмия в зоне нахождения таких нейронов вызывает саккады соответствующего направления.
Таким образом, на нейронах верхних слоев двухолмия имеется полная упорядоченная проекция сетчатки. Нейроны нижних слоев также ретинотопически упорядочены, и их моторные поля совпадают с соответствующими зрительными полями нейронов верхних слоев. Нейроны промежуточных слоев имеют упорядоченные соматические проекции передней части животного (голова, верхние конечности), а также упорядоченные проекции слухового пространства. Все это указывает на большую роль зрительного двухолмия в механизме ориентировочного поведения.
Движения глаз и сенсомоторная интеграция при зрительном восприятии. Глазодвигательная система человека выполняет следующие задачи: 1) сохраняет неподвижным изображение внешнего мира на сетчатке во время движения относительно этого мира; 2) выделяет во внешнем мире некоторые объекты, помещает их в зоне сетчатки с высоким разрешением (зрительная ямка, fovea) и прослеживает их движениями глаз и головы; 3) скачкообразные (саккадические) перемещения взора для сканирования (рассматривания) внешнего мира. Краткие сведения об устройстве периферического звена окуломоторной системы были приведены выше.
Напоминаем, что саккады - это быстрые содружественные отклонения глаз в начальной фазе реакции прослеживания, когда скачком глаза «захватывается» движущаяся зрительная цель, а также при зрительном обследовании внешнего мира.
Дата добавления: 2016-03-05; просмотров: 754;